探究变叶海棠物种界定和居群遗传分化模式
时间:2015-01-24 来源:未知 作者:傻傻地鱼 本文字数:3935字
物种复合体(species complex)是指通过杂交和多倍化产生的亲缘关系复杂、遗传分化很小的多类群复杂物种系统(Koch et al.,2003;Lo et al.,2010;Durkovic et al.,2012)。变叶海棠物种复合体(Malus toringoides species complex)是苹果属野生资源中变异式样和种间关系很复杂的一个复合体,由变叶海棠、花叶海棠、陇东海棠、多毛海棠、马尔康海棠和小金海棠 6 个物种组成(成明昊 等,2000,2002,2003;石胜友 等,2004)。AFLP 标记分析和多拷贝核基因 ITS 序列变异支持变叶海棠是花叶海棠和陇东海棠杂交后代的观点(石胜友 等,2005;Feng et al.,2007);通过详细的野生资源调查研究,另外 3 个与变叶海棠近缘的类群被界定为物种并归入变叶海棠物种复合体。
其中马尔康海棠的花、叶和果实等分类学性状介于变叶海棠和陇东海棠之间,而多毛海棠的性状则介于变叶海棠和花叶海棠之间(成明昊 等,2002,2003;邓洪平 等,2002)。小金海棠的叶片和花部性状与陇东海棠近似,而核型与四倍体变叶海棠相似(梁国鲁和李晓林,1993;成明昊 等,2000)。
在地理分布上,变叶海棠物种复合体目前界定的 6 个物种在四川西部山区呈同域分布(成明昊 等,1999)。上述证据显示这些物种彼此具有非常紧密的亲缘关系。
前人的研究不仅丰富了对野生苹果资源的认识,也在一定程度上揭示了变叶海棠物种复合体中物种的遗传来源(邓洪平 等,2002;石胜友 等,2004)。与多拷贝核基因(如 ITS 序列)相比,单拷贝核基因用于野生果树资源的研究具有如下优势。首先,避免了致同进化(concerted evolution)导致的序列均一化,杂种中两个亲本所特有的核苷酸变异可以完整保留下来,有助于杂种的识别和杂交亲本的确定(Sang,2002)。其次,单拷贝核基因基本不受基因重复的影响,能够很好地反映物种真实的进化关系(Zou et al.,2008)。单拷贝核基因已经成功应用在作物资源遗传关系的研究上(Geet al.,1999;Doyle et al.,2004;Brassac et al.,2012)。使用编码淀粉分支酶的单拷贝核基因 SbeⅠ探究变叶海棠物种复合体中 6 个物种的遗传关系。SbeⅠ基因分析进一步证实变叶海棠源于杂交,花叶海棠是杂交亲本之一,但是排除陇东海棠参与了变叶海棠的杂种起源。同时 SbeⅠ分析显示多毛海棠与变叶海棠,以及马尔康海棠与小金海棠具有相同的遗传组成,并且后两者是变叶海棠与陇东海棠进一步杂交产生的后代(Tang et al.,2014)。
物种分类界定不仅是分类学和进化生物学研究的重要问题(De Queiroz,2007;Fujita et al.,2012),也是野生资源鉴定、评价、利用和保护研究中需要首先解决的基本问题。变叶海棠物种复合体中的类群能否界定为 6 个独立的物种?如果不能全部界定为物种,哪些需要归并?至今没有得到解决。本研究在阐明变叶海棠物种复合体类群遗传来源的基础上,通过单拷贝核基因 SbeⅠ探究其物种界定和居群遗传分化模式,确定复合体类群的分子分化水平,并阐明类群的分类学地位,以期为苹果属野生资源的鉴定、利用和保护提供指导和科学依据。
1、 材料与方法
1.1 变叶海棠物种复合体物种取样
2011 年 6 月在四川省阿坝藏族羌族自治州的阿坝县、马尔康县和小金县采集变叶海棠物种复合体居群样品(表 1)。一共采集了不同地点的 10 个自然居群的叶片材料,每个居群随机挑选 2 ~ 6 个个体,不同个体间隔 30 m 以上,叶片材料经硅胶干燥后带回西南大学,保存在–20 ℃冰箱中备用。
变叶海棠物种复合体除花叶海棠和陇东海棠外,其余均为无融合生殖类群,同一类群仅由一种基因型构成(Tang et al.,2014),因此现有居群取样基本能代表整个变叶海棠物种复合体。
1.2 PCR 扩增、克隆和测序
采用 CTAB 法(Doyle & Doyle,1987)提取叶片总 DNA,稀释至合适浓度。SbeⅠ基因的引物根据栽培苹果的 SbeⅠ基因外显子序列设计,上、下游引物分别为 M1014F(5′ ATAGTGAGCAGCACAAATG 3′)和 M1014R(5′ TGCCTCCTCACTTTCAT 3′)。PCR 反应体系为 25 ?L,包括 2.5 ?L 10×Ex Taq 缓冲液,10 ~ 50 ng 的基因组 DNA,2.0 mmol ? L-1的 MgCl2,0.2 mmol ? L-1的每种 dNTP,0.1?mol ? L-1的上下游引物,1.0 U 的 Ex Taq 酶。扩增程序:94 ℃预变性 1 min,然后 94 ℃变性 30 s,55 ℃退火 30 s,72 ℃延伸 1 min,30 个循环,最后 72 ℃延伸 10 min。扩增产物在 1.5%的琼脂糖凝胶中通过电泳分离,纯化回收之后克隆至质粒载体,转化 TOP10 大肠杆菌。每个样品挑选 8 ~ 16个阳性克隆,提取质粒后送英骏公司(重庆)的 ABI3730 测序仪测序。
1.3 物种界定分析
SbeⅠ核苷酸序列的多序列比对由 ClustalX 软件(Thompson et al.,1997)在默认参数设置下进行,然后手工调整同源性排列不恰当的部分。PCR 扩增过程中产生的重组序列由 RDP3 软件(Martinet al.,2010)识别并去除。挑选代表性 SbeⅠ序列用于后续数据分析。根据 Modeltest 3.7 的 AIC 优化标准(Posada & Crandall,1998)确定 SbeⅠ最优核苷酸替换模型,然后使用 PHYML 软件(Guindonet al.,2010)构建 SbeⅠ最大似然进化树,进化树拓扑结构的可靠性通过 100 次自展(bootstrap)检验进行评价。我们使用 GSI 指数 genealogical sorting inde(xCummings et al.,2008)和 PTP 模型 Poissontree processes(Zhang et al.,2013)在两个进化等级上进行物种界定分析。一是将 SbeⅠ的整个最大似然进化树用于物种界定,同时也对进化树的 3 个进化枝逐一进行物种界定分析。为了考虑拓扑结构估计中存在的不确定性,利用 100 次自展检验得到的进化树构建主要规则一致树,将其用于物种界定分析。所有的界定分析均通过在线分析工具(GSI 指数)和PTP 模型)完成。
1.4 居群遗传学分析
对于 3 个进化枝以及进化枝的每个类群,使用 DNASP 程序(Librado & Rozas,2009)计算 SbeⅠ基因的核苷酸多样性,指标包括分离位点个数、单倍型个数及其多样性、核苷酸多样性(π 和 θ 两种度量指标)。变叶海棠物种复合体分类群间的成对固定指数 Fst也通过 DNASP 程序计算。采用MEGA5.10 程序(Tamura et al.,2011)计算不同类群的成对遗传距离,同时也计算同一类群内部的遗传距离。另外,计算任意两条序列之间的遗传距离。遗传距离根据 MEGA5.10 程序的 Kimura 双参数模型(Kimura,1980)进行度量。
2、 结果与分析
2.1 序列特征
共获得了变叶海棠物种复合体的代表性 SbeⅠ基因序列 82 条,通过多序列比对,经手工调整后长 983 bp,包含 SbeⅠ外显子 11 ~ 17 之间的核苷酸序列。SbeⅠ序列包含变异位点 26 个,其中简约信息位点 22 个,绝大部分变异位点分布在内含子区域。
2.2 变叶海棠物种复合体物种界定结果
Modeltest3.7 确定最适合变叶海棠物种复合体 SbeⅠ基因的核苷酸替换模型为 HKY + I。基于HKY + I 模型使用 PHYML 构建了 SbeⅠ基因的最大似然进化树,结果如图 1 所示。
变叶海棠物种复合体的 SbeⅠ被分为 3 个大的进化枝(Clade 1 ~ 3),每个进化枝作为单系得到了很好的支持,但是进化枝内部的分歧关系基本没有自展支持。进化枝 1 包含变叶海棠、多毛海棠、马尔康海棠和小金海棠 4 个分类群;进化枝 2 由进化枝 1 的 4 个分类群再加花叶海棠构成;进化枝3 由陇东海棠、马尔康海棠和小金海棠组成(表 1)。SbeⅠ基因最大似然分析结果与变叶海棠物种复合体遗传来源研究结果 Tang 等(2014)的一致。
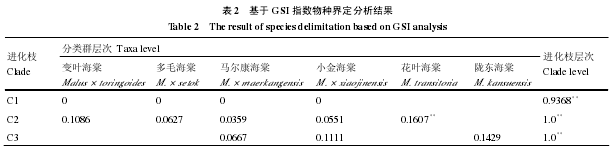
使用 SbeⅠ基因的整个拓扑结构进行物种界定分析,GSI 指数分析显示每个进化枝的 GSI 指数接近或者等于 1,表明这 3 个进化枝可以清晰地界定为物种,并且 P 值在 0.01 水平上显著(表 2)。PTP模型分析的结果和 GSI 指数的相同。
为了进一步探讨组成进化枝的不同类群是否可以界定为独立的物种,对每个进化枝逐一进行了GSI 指数和 PTP 模型分析。GSI 指数分析的结果见表 2,组成进化枝 1 的 4 个类群的 GSI 指数均为0,对应的 P 值全为 1,显示进化枝 1 的 4 个类群不能被界定为物种。组成进化枝 2 的 5 个类群中,仅花叶海棠 GSI 指数的 P 值在 0.01 的水平显著,其他 4 个类群 GSI 指数的 P 值均不显著,表明这 4个类群在进化枝 2 上仍然不能界定为物种。进化枝 3 的 GSI 指数分析显示其所包含的 3 个类群也都不能被界定为物种。虽然陇东海棠 GSI 指数的 P 值不显著,但是 GSI 指数值相对较大,表明陇东海棠 SbeⅠ基因谱系汇聚的程度比马尔康海棠和小金海棠的高。PTP 模型分析与 GSI 指数的物种界定结果基本一致,进化枝中的每个类群作为物种均未得到支持。
2.3 变叶海棠物种复合体的遗传多样性和遗传分化
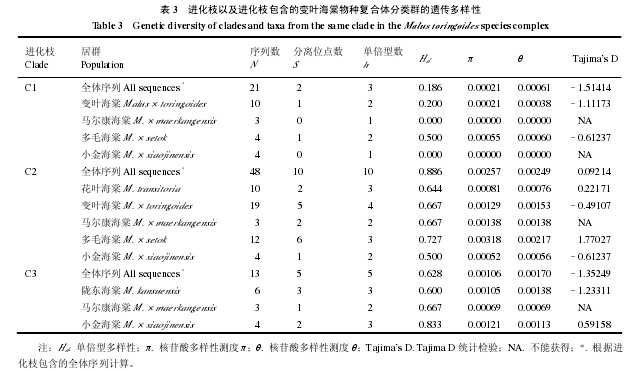
Tajima’s D 检验表明 SbeⅠ基因在变叶海棠物种复合体分歧过程中没有偏离中性进化模式(表3)。由于进化枝之间的遗传分化较大,因此以进化枝以及进化枝包含的分类群为单位计算遗传多样性,结果见表 3。以进化枝为单位,遗传多样性由低到高的顺序是:进化枝 1 < 进化枝 3 < 进化枝2。以分类群为单位,多毛海棠的遗传多样性(π)大约是变叶海棠的 2.5 倍。马尔康海棠和小金海棠的遗传多样性与其所处的进化枝有关。进化枝 1 中两者均无变异,进化枝 2 中马尔康海棠的多样性高于小金海棠,而进化枝 3 中则是小金海棠的多样性高于马尔康海棠。
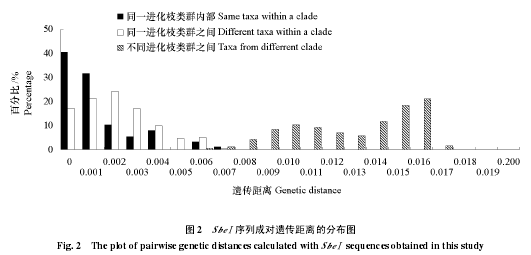
采用遗传距离和固定指数 Fst两个指标度量类群间的遗传分化,结果见表 4。不同进化枝类群间的成对遗传距离基本上都大于0.01,而同一进化枝的不同类群的成对遗传距离大多在0 ~ 0.001之间,仅进化枝 2 的类群彼此的遗传距离在 0.002 ~ 0.004 之间。进一步计算任意两条 SbeⅠ基因序列的成对遗传距离,并绘制遗传距离的分布图,结果见图 2。就同一进化枝而言,类群内的遗传距离和类群间的遗传距离在分布范围上重叠,而位于不同进化枝的类群其成对遗传距离主要分布在 0.009 ~0.016 之间,几乎不与前两者的遗传距离分布重叠。成对遗传距离的分布模式显示,进化枝中不同类群彼此间的遗传分化水平与同一类群不同个体间的遗传分化水平相当。
- 相关内容推荐
- 武都区油橄榄生产施肥问题探究2016-04-28
- 玉铃花种子休眠解除的理化变化探究2016-08-16
- 不同NaCl胁迫下的苹果砧木山定子和八棱海棠的光合特性2014-09-17
- 宁夏设施桃树栽培中的整形修剪问题探究2016-01-15
- 探究红叶李对伴生草坪植物的化感作用2015-01-24
- 探究对涪陵榨菜产量和品质影响较好的施肥方案2014-07-07
- 草莓果实的成熟软化机理探究2015-10-19
- 无花果适宜的贮藏温度探究2015-10-19





