����DNA�о�ʧ�ܵ�ԭ��̽��
ʱ�䣺2014-08-08 ��Դ��δ֪ ���ߣ�ѧ���� ����������10395��
����DNA ��ѧ������������������ѧԺ�ı��֣�Linus Pauling�������Robert Corey����Ӣ�����ĵ���ʵ���ҵ���ɭ��James Watson��������ˣ�Francis Crick��֮����С�1953��2��21�գ����ֺͿ����ڡ���Ȼ����Nature�������о���“����Ľṹ”��Structure of the Nucleic Acids����
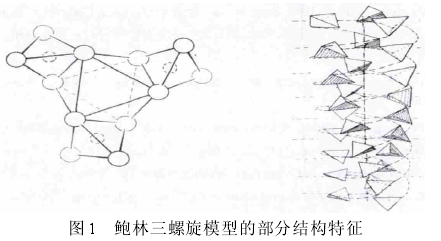
����“����X��������ͼ����Ҫ�����ͻ����ķ��ӽṹԭ������������˺���Ľṹ������ṹ��������������Ƶ�������������� ��������ط��ӵ���������ܶѻ������Dzл�Χ������Χ�����ʺ�����������ſ���ƽ��������ᴹֱ��”��[1], p.346��1953 ��������ֺͿ������ڡ�������ѧԺԺ������Proceedings of National Academy of Sciences������“Ϊ��������Ľṹ��”A Proposed Structure for the Nucleic Acids������ϸ���������ʼ�����़�λ�ڷ��ӵ���࣬����������������֮��������ϵ���������ģ�͡�ͼ1��Ϊ�����ḽ��������������������ţ�ʵ��ԲȦ������ԭ�ӣ�����ԲȦ������ԭ�ӣ���ͼΪ24-�л�7-ת�ĵ������������������ʾ������ţ�ૺ������������ǵĻ��߱�ʾ����[2], pp.87-88����
����
����1953 �� 4 �� 25 �գ���ƪ DNA �ṹ���������������ڡ���Ȼ���ϣ�����������˹��Maurice Wilkins����˹�пˣ�Alex Stokes��������ѷ��Herbert Wilson����“�������Ǻ���ķ��ӽṹ”��Molecular Structure ofDeoxypentose Nucleic Acids�����������֣�Rosalind Franklin������˹�֣�Raymond Gosling����“���ٺ����Ƶķ��ӹ���”��Molecular Configuration in Sodium Thymonucleate������ɭ������˵�“����ķ��ӽṹ”��Molecular Structure of Nucleic Acids������[3], p.737����“����ķ��ӽṹ”�У���ɭ�Ϳ���������DNA ��˫�����ṹ����������ƽ�еĶ��������Χ��ͬһ����������ƣ���������़�λ��˫�������ڲ࣬�������������������֮��������ϡ�1953��5��30�գ��������ڡ���Ȼ������“�������Ǻ���ṹ���Ŵ�����”��Genetical Implications of the Structure of Deoxyribonucleic Acid������һ���ش���“�������Ǻ���������Ҹ���”�����⡣��ɭ�Ϳ����Ҳ��˻��1962���ŵ��������ѧ��ҽѧ��������DNA ��ѧ����������Ϊ“Զ������Ĵ���ʿ����������С�俳��”��[4], p.436���Ĺ��£�20 ���ͽܳ��Ļ�ѧ�ұ�����“����������”�������˴�ܵ�ԭ��ֵ������й���ɭ�Ϳ���˷���DNA˫�����ṹ�����źܶ࣬�����ڱ���DNA�о�ʧ�ܵ�ԭ����δ����صı�������������һЩ������̽�֡�
����һ������ʧ��ԭ�����Ҫ�۵�
��������DNA�о�ʧ�ܵ�ԭ����ʲô��������һ���⣬��ν���ʼ��ǣ�������Թ���Ϊ���¼��ֹ۵㣺
����1.“����ȱ��”˵
����X ��������ͼ�� DNA �о��Ĺؼ�֮һ��20 ���� 50 ������������� DNA ��������ͼ���ѻ�ã���Ҫ����DNA������ճ���ĵ����ʷ��룬���ҷ��������Ҳֻ������ͼ�dz�ģ����DNA���Ρ���Ȼ����ѧԺ���ʵ¶���John Randall��ʵ�����Ѿ��������������X��������ͼ�����DZ��ֺͿ���ʹ�õ���Ȼ��Ӣ������ѧ�Ұ�˹�ر��William T. Astbury��1938�귢����“�൱ģ��”��X��������ͼ�������ڡ��ṹ��ѧ���������ѧ��ʮ��Ľ�չ����Fifty Years of Progress in Structural Chemistry and Molecular Biology����ָ����“������Щ�൱ģ����X������Ƭ����ʵ����Ӧ��DNA��ά��������ʽͼ�ε��ص������ǵ����ҵó������Ľ��ۣ�DNA��ά���������ӱ˴����Ƶ��������������������������и��ݻ����۾����������������˫������”
����1951 �����죬�������Ӽ�������ѧԺ�������ڼ������� · ��˹�أ�Gerald Oster���Ľ����º��ʵ¶���ϣ����һ��������˹��X��������ͼ��1951��8��28�գ��ʵ¶��ڻ�����д����“�ܱ�Ǹ����˹�������������DNA�������ͼ��������˹����������æ�ڽ���X��������ͼ������Щ��Ƭ���������������ǻ�������ʵ���ҵ�Ŭ����˵���Dz���ƽ�ġ�”
����1952��5�£�����ԭ�ƻ���ϯӢ���ʼ�ѧ��ĵ����ʻ��飬���ҷ����ʵ¶���ʵ���ң���ʱ�������������B��DNA������ά��X��������ͼ���������ʾ��˫�ضԳ��ԣ������������ų����������Ŀ����ԡ��ڻ����ǰϦ����Ϊ“�������е����в������������������”�����������ܾ�������ǩ���������ա���ʷѧ������·�¶�����Robert Olby����Ϊ��������ֵ��Գ��У�“������˹�ǻ��ó���Щ������������”����[5], p.49���������ֺ��п��ܽ�ʾDNA����ȷ�ṹ��
����2.“����”��“�Ը�”˵
�������ֵĴ�����������˹·����Thomas Hager����Ϊ��“����”��“�Ը�”�DZ���DNA�о�ʧ�ܵ���Ҫԭ��֮һ��20 ���� 50 ������DZ��ֿ�ѧ���ĵ�һ���߷塣1951 �� 4 �£����ֺͿ����ڡ�������ѧԺԺ��������“�����ʵĽṹ����������������������”����[6], p.205���������ļ���ƽ����Ϊ�������ɹ����ÿȦ3.6��������л���������Ȧ֮���γ����������α-�����ṹ��֮���ֺͿ����ַ���“�������������������͵�ԭ������ͽṹ����”��“�ϳɶ������Ľṹ”��“ë���������������ص����ʵĽṹ”��“Ѫ�쵰�������Ķ���������”����ƪ���ģ�“Ϸ���Ե�”ռ�ݡ�������ѧԺԺ����1951��5�ºŵ�ȫ�����档
�������Ŵ�ѧ�Ŀ��ĵ���ʵ����Ҳ�������ѧ������˹·������William Lawrence Bragg�����쵼�½��е����ʽṹ�о���1950��3��31�գ������ϵ�³����³����Ӣ�����ʼ�ѧ��ѧ������Proceedingof the Royal Academy����������“���嵰�Ķ���������”��Polypeptide Chain Configurations in CrystallineProteins�����뱫���෴�����ĵ����о�С�鷸��������Ҫ�Ĵ���һ�Ǿ����������������ϸ���ÿһȦ�İ�����л���ĿΪ3��4�����������Ǻ������ļ���ƽ���ԣ�û�дӻ�ѧ���Է���ʩ�����ƣ���������ĵ����ʽṹ�ﵽ��ʮ��֮�ࡣ
����սʤ������������ѧ�Ҹ����ִ����˼����������Ҳ��һ���̶��ϲ�����“�Ը�”���������������Ϊ��“�ڽ���˰������������������Ϊ�Լ�������Ҫ��������Ҫ������Щ�о����������ˡ�����������������Ͻ������ӽṹ�������ѡ��”��[4], p.435��α-�����о�ǰ����ʱʮ���꣬�����о�С����������ӵļ����ͼ��Ƿ������˴����Ĺ�������DNA�о�ȴʮ�ֲִ١��������������ıȽϣ�“��������������ʮ���귭����ȥ���з����Ľ�����������������ڳ�ǧ����Сʱ�м��ϸ�µؽ��о��������о�����Ѫ��������ʽ��������ģ��ǰ������ʵ���ҽ�������ĸ����ɷ־�ȷ�زⶨ���˼���֮һ�Ȼ�ٷ�֮һ����
�����ڵ�������һ�����ϣ���������X�������㵽�Ĵ������������ϣ������ñ�������ϸ���о���ʹ���п��ܽ���ʮ�ֲ�ͬ�Ľṹ���� ����DNA����Щ���鶼û������”��[4], pp.427-428��
3.“�Ͽ�ѧ��”֢״
������³�ģ�Max F.Perutz�����о�Ѫ�쵰�Ľṹ�����1962��ŵ������ѧ�������ڡ���������ŭ��—���ڿ�ѧ����ѧ�Һ����Ե���ʡ���I Wish I'd Made You Angry Earlier-Essays on Science, Scientists, andHumanity����ָ����“ֱ�����������һϢ�������Բ�ͣ�ط������ģ�����Щ����û��һƪ�����ڵ�����������Ҫ��������Ϊ������רע��ijЩ���飬�����Ǻ�ս�����Ժ�����ά����C�����⡣”��[7], p.13������DNA�о�ʧ�ܺ���ʡ����ѧԺ������ѧ��Ҳ���ƲⱫ���Ƿ���“�Ͽ�ѧ��”֢״�����ǰ�����֢״����Ϊ“ΰ�����������˿�ѧ������ѶԷ����������ϵͳʱ�������ġ�”��[8], p.50����Щ�۵��漰����ѧ�ҵĴ����ԺͿ�ѧ�о��ķ�������ѧ�ҵĴ����Գɾ�������������ʱ�������ģ�“����”��ѧ����֪ʶ�;�������ƣ����ǣ�����Ҳ������˼ά��ʽ��Ӱ�죬Ҳ������Ϊ�������������������������������������ǿ˷����˾�����Ч�������ڱ��ֵ�ʵ���ң���һ��“��ʦ���о������������ĵ�λ�������Ȩ����������”��[4], p.187�����о���������о�������ڷ��Ӽ�����ǻۣ������ڿ˷�������ʶ�ľ��ޡ���֮������DNA�о�ʧ�ܵ�ԭ���Ƕ��ģ�����ÿһ�ֹ۵㶼��һ�����������ǻ����Խ�Ͼ����о���������һ���ķ�����
����������ʶ�ͷ����ľ����ǵ��±���ʧ�ܵ�����
����1. ��“�Ŵ���Ϣ������”����ʶ�ľ���
�����κο�ѧͻ�ƶ���Ҫ�����Ļ��ۣ�������տ�ѧ�����ı仯����������ѧ���ֵ���Ҫǰ�ᡣ20����50 ���������ѧ��չ�Ѿ�Ϊ��ʾ DNA �Ľṹ��������������Ъ����Friedrich Miescher������������AlbrechtKossel�������ģ�Phoebus Levene���Ѿ���ʶ���� DNA ����ɡ���������ӷ�ʽ��������ѧ�����ġ�Ӣ������ѧ�Ұ�˹�ر��Ų������ѧ�Ҹ��������Ⱥ������DNA�Ľṹģ�ͣ�X������������ҲΪ��ʾDNA �ṹ����ʵ������������Ҫ���ǣ�Խ��Խ���֤����ʾ DNA �����Ŵ���Ϣ�����塣���ǣ����ֶ���DNA�������ʶȴ������һ�����۵Ĺ��̣�
������1��20����40�����������Ҳ“��ʧ”�ڵ����ʵ�ƫ���С������ĵ��ĺ������˵�У����ּ����DNA�еĺ�����ȣ��������ظ��ṹ��DNA�ƺ����ʺ�Я���Ŵ���Ϣ������������20�ֲ�ͬ�İ�������ɣ����Ǿ��ж�����ʽ���ӵ����з�ʽ�������Խṹ�������ʺϴ��渴�ӵ��Ŵ���Ϣ���뵱ʱ���ձ���ʶһ��������Ҳ��Ϊ�ṹ��DNA�ܿ���ֻ��һ��“�ṹ��”�ijɷ֣���������������Ŵ���Ϣ�����塣
������2��ʵ���ϣ�19����60�����Ъ�����о��Ѿ�ԤʾDNA���Ŵ����壬�������Ĺ���“�ںܴ�̶���”�������ˡ�ֱ��1935�꣬��˷����о�Ժ�İ����Oswald T. Avery�����֣�DNA���������ڷ������֮�䴫���µ��Ŵ���Ϣ�����������͵¡��ٵ�ϸ������ת���о���ʾ���Ŵ���λ�������ڸ��ӵĵ������У��������ڱ��濴����������DNA�С�1944�꣬����������ģ����DNA�ڷ������֮�䴫���Ŵ����ԵĹ۵㣬���ҽ����˸����˹��“ת������”����������������ڡ�ʵ��ҽѧ��־����Journalof Experimental Medicine���Ϸ���ʱ�����ֲ�û�������ı���� DNA ����Ŀ�������˵��“����ǰ��֪��DNA ��һ���Ŵ����ʵ��۵㣬Ȼ������û�н�����һ�۵㣬����֪������ʱ���������ڵ����ʵ��о�������Ϊ���������п������Ŵ����ʣ��������Ǻ���—��Ȼ������Ҳ�����á�”��[4], p.400����3��������ʶ��DNA�������Ƿ���“¬�����ɾ������”֮��1952��7�£����ֳ�ϯ�ڷ������еĹ������ﻯѧ��ᣬ֮���ֲμ��ڰ���ǽ���¬������Ժ���еĹ����ɾ�����飬��ȡ��������ѧ�Һն�ϣ��Alfred D. Hershey���ı��档1952�괺���ն�ϣ�Ͳ�˹��Martha Chase�����÷�����ͬλ���ף�P32������S35����T2�ɾ����DNA��ǣ����Ƿ��֣����ɾ�����Ⱦ�����ϸ��ϸ��ʱ��ֻ���ɾ����DNA����ϸ���ڲ�����������ȴ�������棬ʵ����ȷ���֤�ݱ���DNA���Ŵ���Ϣ�����塣“¬�����ɾ������”֮���ֲ���ʶ���Լ�“�����˴���ķ���”��
���������˵��“˫�����ķ��ֹ��̣��ӿ�ѧ��˵����ʮ��ƽ���ģ�����Ϊ��һ���DZ������ǿ���ġ���Ҫ�� ���ڷ�����DNA�ṹ������Ȿ����”��[9], p.55���ֵ�ɭ��Horace Judson��Ҳָ����“����DNA�ṹ�ڵ�ʱ�Ǻ��ѵģ����������������ڵ�ԭ������Ϊ����û����ʶ��������Ҫ�Ժ������ԡ�”��[8], p.8�����ȣ�DNA������“�������Ҫ��”���������������о�����Ҫ���壬���Ҷ���ȷ�о�DNA�ĽṹҲ������Ҫ��ָ�����塣���ڱ�����˵����ֻ�ǰ�DNA������һ�ֱȽ���Ҫ���������Ӷ��ѣ�“����ֻ���������˸���Ȥ�Ļ�ѧƷ�������Ȼ�����һ����Ȥ�Ļ�����һ����”��[4], p.434����ʶ�ľ����������о��ߵ���Ұ���뱫�ֲ�ͬ����ɭ�Ϳ���˸������ʶ����DNA�����壬���ҽ��о�Ŀ��ȷ��Ϊ“����������ʲô”����“�ڽ�ʽ��Ŀ��”�������Ǵ�һ��ʼʱ�;��и��ӿ����о���Ұ����Ҫԭ����Σ���ʱ�����������֡��������ɭ������˶��Ǵ�1951���������ʼ�о�����1953��4����ɭ�Ϳ�����ڡ���Ȼ������“����ķ��ӽṹ”Ϊֹ��ǰ������ʱ�䡣��ȷ��ʶ�о���������壬�����Լ��������Ͷ�뵽�о���ȥ������һ�������Ե���Ϊ��Ҫ��
����2. ���о����̿�˼ά��ʽ�ľ���
�������о������������ܵ�����˼ά��ʽ��Ӱ�죬���DZ���ʧ�ܵ���Ҫԭ��������Ժʿ1946��9�µ�1951���ڼ�������ѧԺѧϰ�����汫������X���߾���ѧ������“��ѧ�Ļع˺�չ��”��д����“����ͨ��ģ�ͷ����ɹ��Ľ�ʾ�˵����ʵ�α-�����ṹ���ڵ����ʽṹ��DNA�Ľṹ֮����ڵIJ���ǣ���Ϊһ���������δ��һ��α-����ʱ��ֻ�迼��һ��������������γɵĵ�������20 �ְ�����л��Ķ��� R ������������࣬�Ӷ�����Ӱ����������Ĺ����� �� α-����������½�������л���R���������������������ʹ���������屣�ֹ����ԣ����������Ϊ�����뷨�ܿ������ڴDNA������ʱ����ؽ�˳���ò�����4�ֲ�ͬ���л��������ಢ������������ڲ�ĵ���”��[10], p.4��1951 �� 11 �£����ִӡ�������ѧ��ѧ������Journal of the American Chemical Society���������»� ·���£�Edward Ronwin��������“������������ṹ”��A Phosphor-tri-anhydride Formule for the NucleicAcids���������µĺ���ṹ�У����Ӻ���������������������ÿ����ԭ���� 5 ����ԭ���������������������ܡ���Ȼ����д����������ѧ��ѧ������ʾ“����5����ԭ��ͬ1����ԭ�ӽ����һ�������ܵĽṹ�����������������ṹ��Ӧ��̫���档”��[11], p.378����ȴ��ͬ���½�������ڷ��������뷨�����絰������һ�����Ѱ�������ڷ��ӵ����Ͳ���Ӱ������Ĺ����ԣ���DNA�У���˫�������ʼ�͵�������़���ڷ��ӵ��ڲ࣬�ʹ�����η��õ����⣬�������Ƿ��ڷ��ӵ���࣬�ͱȽ�����������ӵ�ģ�ͣ����ң���˹�ر����X��������ͼ��û���ų����ֿ����ԡ���˱����Ʋ⣬DNA�еļ���ܿ����ڷ��ӵ���ࡣ
�����ֻ��һ�ֳ����ļ��裬�����������о��н��м��飬Ҳ���뿼�ǵ����������ԡ����DZ��ֽ�������ڷ��ӵ�����“һ������������İ�װ”����[4], p.434�����ڶ����������“ƫ��”������û�����濼�������˲�ͬ���о��ɹ����ڼ����Է�ʽ�ϣ�����˵“����Ȼ��֪��E.��٤��Ĺ���������ָ������DNA�У����ʺ���़���Ա�����ȣ�������ȴû���뵽������ϵĻ����ṹǣ�浽��������������ऺ�������������֮���������⡣”��[5], p.50����˫�������ϣ�����˵��“����Ϊ���������1953��2���������������ṹ�ܿ����Dz�������ά�γɹ��̶�����������״̬�²���ʾ��DNA���ӵ��˹���Ʒ��”
����˼ά��ʽҲ������DNA�ṹì�ܵĴ����ϡ���α-����һ����������ģ��Ҳ��“���ì�ܵ�����”��[4],p.424�������о������20 ���� 30 ��������־��Ѿ���������ʵij����ṹ���������ֽṹ�뵱ʱ�ľ���ѧȨ����˹�ر����X��������ͼ������ì�ܣ��ڰ�˹�ر����X��������ͼ�ϣ���������ÿ��5.1����������ظ���λ��������ģ����һ������5.4������������ģ�͵Ĺؼ��ṹ֮���ִس�����һ���ݵ����������ѧ�繫����α-����ģ�͡�ͬ������DNA�ṹ�У���Ȼ����ļ��������DNA���IJ�λ��������̫������“װ��”��һ�𣬵��DZ�������ؿ������ڷ��ӵĺ��IJ�λ��ÿ��ƽ����������������ţ�ÿ����������������ṩ2����ԭ�ӣ�����һ�������İ�������״����[2], p.89�����Ǿ���ѧ�еij�����״���ƺ������ṹ��“������”�����ڵ����ʵľ��飬����ͬ������Ҳ��“�ں������ҵ���Ҫ����ʵ���ش�����������”����[4], p.425��һ���棬���ڼ�˵�ļᶨ�������α-�����ijɹ�����һ���棬����˼ά��ʽȴʹDNA���о�“����������ͬ”���ܽᱫ����DNA�о��е�ʧ������Ϊ��
������1����ʶ��Ƭ���ԡ�������⣬����������Щ“�Զ���”�Ľ��ۣ�û�о����ϸ�ļ����ȷ��������������Լ�����һ�����ķ����У�����²�ŵ��Edward Bono����ָ���ģ�“������ڴ��˵ط�����ô�ٶ�ĸĽ�Ҳ����ʹ����λ����ȷ��”��[12], p.14��
��2���ڿ�ѧ�о��У�“ƫ��”������Ӱ���о��߿۵Ĺ۲���жϣ�“����Ը��”�ڲ�֪�����в�����ʵ���ϣ���٤�����˫������ʵ�����Ѿ�֪��������“ƫ��”ʹ�о���û�����������ɼ�˵��ͬʱҲ�������Ծ��������˶���ʵ���жϡ��������棨William Beveridge��˵����“һ��������ij������֮�ͺ��ѽ������ڷ������෴����ʵ����������”��[13], p.55��“���DZ���ʮ��ע�⣬��ʹ�Լ����Լ��ļ�˵�������У�Ӧ����۵��ж� Ҫ��߾��裬��ʹ�۲�ͽ����ܵ���˵��Ӱ���������”��[13], p.55����ɭ�Ϳ����Ҳ������������ʶ�����˵�������֡���ŵ�ݣ�Jerry Donohue������������ɭ�Ϳ���˿˷�������ʶ��“����”����ô��“����”������Ҳ����������Ը�����������������1951��11�£���ɭ�Ϳ���˸���X��������ͼ��������С�ͺ�ˮ����������һ���뱫�����Ƶ�������������-���������ڷ��ӵ��ڲ࣬����ڷ��ӵ���࣬��������������������þ�����γɵ��μ�ά���������ȶ�����ɭ��Ϊ����������pH��������������磬һ�����������ӵ�������ĸ���ɣ�“���þ���ӻ�����Ӹ����Ǻ�����ĹǼ�֮�У��ܿ���ܵõ�һ�ַdz����µĽṹ������ȷ���ɷ��顣”
����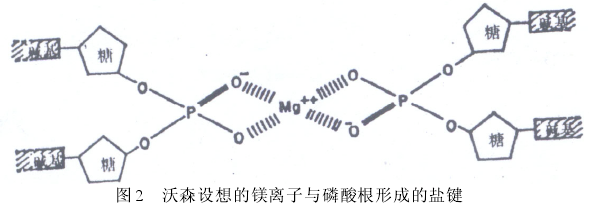
����Ȼ������һ“��ȷ���ɷ���”������ܿ�ͱ��������ַ��ˡ�1951��12�£����������������ţ�ֱ���˵���ָ�����ǵ��������ԵĴ���һ����ϸ�����У�þ���ӻ�����ӱ�ˮ���Ӱ�Χ�������ܽ���������һ�𣻵ڶ���DNA�ĺ�ˮ��ʵ����Ӧ�������Ǽ���ֵ��10�������ڸ������ֵ���������ɭ�Ϳ���˼�ʱ��ʶ����Щ��������“ͪʽ”��“ϩ��ʽ”�칹��DNA�ṹ����һ���ؼ�����ʱ�Ľ̿�����Ϊ��ԭ�ӿ��������ʻ�����ऻ��ĵ�ԭ�Ӻ���ԭ��֮���ƶ������ڻ����칹ƽ��֮�У��������Ϊ���ֱ仯�Ĺ��������γ��ȶ��������������������ϣ���ɭҲ��Ϊͬ����ԭ����ΪӦ�ð���-�����������ڷ��ӵ��ڲࡣ����������ѧ�Ҷ�ŵ�ݵİ����£���ɭ�Ϳ������ʶ���������ʻ������������ͪʽ��������ϩ��ʽ����ʽ���ڣ����ڿ˷��˼�������ϵ���Ҫ�ϰ���
������ɭ�Ϳ���˵����Ӵ��෴�ĽǶ�˵���˱��ֵľ����ԣ���β��ܼ�ʱ��“������”ת�䵽“����ڲ�”����“����”ת�䵽“˫��”������ͬ����Ҫ�����о�С�����������������������������ڡ����ȵ���—��ѧ����֮�Ҽ�����What Mad Pursuit-A Personal View of Scientific Discovery����ָ����“��ô���Һ���ɭ����ʲôֵ�óƵ�֮���أ�����еĻ���Ҳ���ǵ�ijЩ����վ��ס��ʱ�������������ڲ�������������� ���ೢ�Ե�ʧ�ܲ�����Ϊ�о��߲�������������������������ͬ���������Ѿͺܿ�����ˡ�”��[15], P.83�����Ҳ��Ҫ�����о�С�����������Եؽ��ܲ�ͬ�Ĺ۵㣬�Կ��ŵ���̬�Դ���٤������Լ�������ģ�͵IJ�ͬ������������ܿ˷�˼ά��ʽ��Ӱ�죬����“��������ͬ”����֮����DNA���о��У�������������Ĺ�����õ�˼ά��ʽ��֪ʶ��“��������”������Ҫ�����籴��������˵�ģ�
����“Ϊ��������ѧ���֣�‘ð�ա��߸����������Լ������˵Ļ���’Ҫ��‘�����˹㲩�����ŵ��ǻ�’��Ҫ�öࡣ”��[16], p.135��“һ��˵������ѧ�����������ֵ�����Ҫ���ء����ع�ʽ����һ����ͻῴ�� ���õ������ͽ����о�ʱ����ȷ��ʽ��֪ʶ�Ĵ������۸�����Ҫ��”��[16], p.133��
3. ������ģ�Ϳ��о������ľ���
�������ֺͿ������������ģ����������Ҫ���⣺һ������ģ�Ͳ��ܽ���DNA�ĸ��ƻ��ƣ���������������������Ӽ�������ϣ���������λ��“��װ”̫���ˡ��ֵ�ɭ�ڡ������͵ĵڰ��졷��The EighthDay of Creation������ָ����
����“��‘�����ɿ��Ľ�����װ�DZ�Ҫ��’��仰��������һ������ʵ—���ְѺ�����װ��̫���� �����Լ���Ь�ӡ���������һ����̫���˻�ʹ�˲���� ģ���Ǽ�Ĭ�ġ���ʲôҲ���ܽ��͡�����û���ṩ�������Ƶ�������Ҳû�б�������ϸ������������ʲôӰ�졣�ر��ǣ��ṹ�����˲�٤���������ģ�͵��κβ��ֶ�û�нҿ���������ܡ�”��[8], p.98����Щ�ṹ����ֱ����DNA������ѧ������㣣���ɭ����Ϊ������֮����û�п��ǵ������������������Ϊ����û��������ѧ����������DNA��ʵ���ϣ�1940�꣬���ֺ͵¶������ˣ�Max Delbrück�����ڡ���ѧ����Science���������ĸ��ƻ��ƣ�������Ϊ������ķ��ѹ��̰��������Σ�����A����Ϊ�ϳɻ�������A—1��ģ�棬Ȼ��A—1����Ϊ�ϳɻ������ӵ�ģ�档���ǣ���ʵ���о��У����ܹ��ڿ���“����Ľṹ��ѧ����”������ǿ����������ý��������˴���ķ������������������µ��룬���ڱ��ֵ����������У��ǻ��������в��ܵ��룬�������ܽ���������һ����������ɭ˵ΰ��Ļ�ѧ�������˻�����ѧ��
����DNA �о����Ա���Ϊ��DNA ���ӽṹ�Ĺ����Լ��仯ѧ�ȶ��ԣ�DNA ���ӵĺ�����������η�ӳDNA �ĸ��ƻ��ƣ�ǰ����Ҫ��“�ṹ”���⣬��������Ҫ��“����”���⡣��������ڷ��ӵ���࣬��Ȼ���ÿ���“��������װ”�����⣬���Ǽ���˴�֮�以����ɣ��������DNA��ȷ�ĸ��ƻ��ơ�����������ڷ��ӵ��ڲ࣬�ڼ�����ӵ���Ϸ�ʽ���������̺��ž�ȷ�ĸ��ƻ��ơ����º���۹�������һ�����ϵ������ʣ�A, Adenine������һ�����ϵ�������ण�T, Thymine���������ʣ�G, Guanine)�����ण�C, Cytosine)����ԣ�������ѧ�ĽǶ�������ÿ������������Ϊһ����ȫ��ͬ���������Ļ������ܹ��ܺõؽ����Դ�������������ͻ���Ȼ�ؿ���“ͬ��”�����“����”������⣬���������ͬ����ԣ����������������ʡ��������������ʡ����������������ड�����������व���ԣ��ͻ�������ͬ������״����ͬ���δ�С�ļ���ԣ�ʹ˫�����ĹǼܲ������ڷ��ӽṹ���ƻ����������涼�������⡣
����������û�����ԣ���A—T��G—C��ԣ���ͼ3������������˷��ӹǼܲ���������⣬���Ҽ��֮�����γɺ��ʵ������Ҳ�̺�������ĸ��ƻ��ƣ������������˽ṹ�빦�ܵ�ͳһ����Ҳ����ɭ�Ϳ�������о���ʱ��ʼ��“��ϵ��DNA�Ĺ���”�����ǿ�ʼ��Ƶľ���“һ���������X������������һ�£�����Ҳ���˼����Դ������˫�ع���”��[9], p.55���ĺ���֮����
������ˣ�DNA�ľ�ȷ��������ͬ����Ի��ǻ��������һ�ṹ���⣬����Ļ�����Բ����ǽ��DNA���IJ�λ�����ԵĹؼ�������Ҳ�ǻش�DNA��ȷ���ƻ��ƵĹؼ������ٿ���˵��DNA�о���������“�ṹ”���⣬ͬʱҲ��“����”���⣬������ʼ���ն�û���漰�������Է�ʽ��һ���̶�����ṹѧ�Һ���������ѧ�����о��йء����չ����˵����“�ڱ��ֵ�ͷ���У������������ڱ����Ͽ��Թ��Ϊ�漰������ṹ��ѧ�������⡣���룬DNA������ѧ�����Ժ�һ����������Ҫ�ǽṹ����ȷ�ģ���ô����ѧ�ϵ���Ҫ����Ȼ��ͨ��ij�ַ�ʽ���ֳ���������һ���ϣ�����������Ū����ṹ����������ȷ�书�ܡ�”
��������ͬΪ“�ṹѧ��”��������˹����ɭָ��“��������˹����ҪĿ�����о����ӱ����Ľṹ������û�д��������Ľṹ�����п���ͨ���κ�ʵ�ʵ�;��ȥ˵������Ŵ����Ĺ��ܡ�������˹��������Ҫ��һ���ṹ���ߡ�”��[17], p.31�����ֵ�ģ�ͷ����������������ͼ��ǵ�“����”��������ѧ������С˼���ķ�Χ��ʹ�о��߿��ԶԽ�����������IJ²⣬���DZ����о��������ŵ㡣���ǣ�������DNA�ṹ�о��У�������DNA������ѧ���ܣ����о������Ͻض���һ����Ҫ��“����Դ”�������Ҳ�DZ���С��ʧ�ܵ�ԭ��֮һ��
���������ᡡ����
����1954 �꣬������ۼ����۵������ŵ������ѧ����1962 �꣬�������Ժ����鵥�����ŵ������ƽ����������ȷ��ʾDNA�Ľṹ�����־ͽ���Ϊ��ѧʷ�ϵ�һ�����λ��ŵ���������ߡ�������������λ��ᡣ���ͬʱ������DNA�������ȷ��ʶ�����õ�˼ά��ʽ��ǡ���о�������������ϣ����ճɾ�����ɭ�Ϳ����“��������ѧ֮��”��������
������Ϊ20����“�ṹѧ��”�Ĵ���������ִ����Ե�����ģ�ͷ������о�������ӵĽṹ���ڿ�ѧʷ�ϳ��ֳ�����-�����벼����-�ϵ�³-��³��֮��ĵ����ʿ�ѧ����������-��������ɭ-�����֮���DNA��ѧ��������“��Ϣѧ��”�Ĵ�ʼ�˲�����Ѧ���̲�ͬ�����ֽ���������һ��“���ӹ���”����Щ˼��������“�������̷��Ӽ��������ı���”��1940�귢���ڡ���ѧ����“�����������������ı���”��1948��Ӣ���ʼ�ѧ���������佲������“���ӽṹ����������”��1948�����·���ػ�����ݽ����У�һ���̶��ϣ�����-��������ɭ-�����DNA�о������IJ����Դ��“�ṹѧ��”��“��Ϣѧ��”�����۵IJ��졣��ˣ��Ա���Ϊ��ע�����ģ������о���صİ�������������20�����ⳡ����Ҫ�Ŀ�ѧ�����Լ���ѧ�����۶������������ʾ������
�����۲� �� �� �ף�
����[1]Pauling, L., Corey, R., Structure of the Nucleic Acids[J]. Nature, 1953, 171(4347): 346.
����[2]Pauling, L., Corey, R., A Proposed Structure for the Nucleic Acids[J]. Proceedings of National Academy of Sciences of theUnited States of America, 1953, 39(2): 84-97.
����[3]Watson, J. D., Crick, F. H. C., Molecular Structure of Nucleic Acids[J]. Nature, 1953, 171(4356): 737.
����[4]����˹·������—20���͵Ŀ�ѧ�ֽ�[M]��������������塢�������룬�Ϻ���������ѧ�����磬1999��187-436��
����[5]L.���֣��ṹ��ѧ���������ѧ��ʮ��Ľ�չ[J]�������ס������룬��ѧʷ��ԣ�1981����1����49-50��
����[6]Pauling, L., Corey, R., Branson, H. R., The Structure of Proteins:Two Hydrogen-bonded Helical Configurations of thePolypeptide Chain[J]. Proceedings of National Academy of Sciences of the United States of America, 1951, 37(4): 205.
����[7]����˹·F.��³�ģ���������ŭ��—���ڿ�ѧ����ѧ�Һ����Ե����[M]���Ŵ����룬�Ϻ����Ϻ���ѧ���������磬2004��216��
����[8]����˹·�ֵ�ɭ�������͵ĵڰ���[M]���������룬�Ϻ����Ϻ���ѧ���������磬2005��8-98��
����[9]���ѣ�DNA���ӽṹ���ֵİ����о�[J]����ѧ�о���1984����7����55��
����[10]����������ѧ�Ļع˺�չ��[J]�������ѧ��2003����4����4��
����[11]Olby, R., The Path to the Double Helix[M]. London: The Macmillan Press, 1974, 378.
����[12]���»�·�²�ŵ������˼ά��[M]��Ǯ���룬����������.����.��֪������꣬1991��14��
����[13]W. I. B.�������棺��ѧ�о�������[M]���½��룬��������ѧ�����磬1979��55��
����[14]ղķ˹·��ɭ��˫����—����DNA�ṹ�ĸ��˾���[M]����룬����������.����.��֪������꣬2001��70��
����[15]������˹·����ˣ����ȵ���—��ѧ����֮�Ҽ�[M]��������Т���룬�Ϸʣ��й���ѧ������ѧ�����磬1994��83��
����[16]W. I. B.�������棺���ֵ�����[M]�������ס����Ƕ��룬��������ѧ�����磬1987��133-135��
����[17]����·E.���ף���������ѧ����Դ�ͷ�չ[J]���������룬��ѧ����ѧ��1982����5����31��
- ��������Ƽ�
- ��ѧ�����Զ���·��������ȱ�������·��2014-07-29
- �ӿ�ѧʵ���г����������ѧ2014-07-28
- ���������ѧ��ѧ���ں������Ӽ�����2014-07-28
- �Կ�ѧ������ѧ��α��ѧ���߹�ϵ������2014-11-15
- ����Ļ�е��·��ǻ�е��·2014-07-28
- �й���ѧ��ѧ�ķ�չ��ʷ����״�����ٵ���ս2015-07-07
- ��ѧ������չ�����ֵ���Ȼ��֤2014-11-15
- ���Թ�ϣ�������Ķ��ֿ�ѧ�Ļ���ѧ��չ������2014-07-29
������ࣺ
�Ƽ��Ķ�





