3 结果与分析。
3.1 低温胁迫对红叶杨和竹柳细胞膜透性的影响。
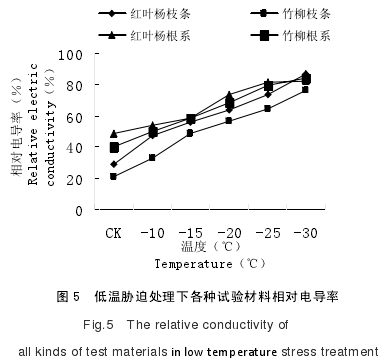
在持续低温胁迫的影响下,植物细胞膜的选择透性会逐渐减弱,从而出现膜内大量溶质外渗的现象。植物细胞膜的损伤程度可通过细胞内电解质渗出的状况判断,并通过相对电导率值的变化来反映[96].植物在经过低温伤害后其细胞膜结构以及功能受到损伤,从而出现植物细胞内电解质不断外渗的现象,致使电导率逐渐升高。根据试验数据显示,随着温度的逐渐降低,红叶杨和竹柳的枝条以及根系的相对电导率逐渐增加,这是细胞膜受到冻害的表现。低温胁迫过程电导率变化如图 5 所示:红叶杨和竹柳的离体枝条和根系的相对电导率随着处理温度的逐渐下降呈现出逐渐上升的变化趋势。相对电导率值变大,反映其细胞膜透过性逐渐增加。
当植物细胞的电解质外渗率为 50%时的温度被称作半致死温度,如图 5 所示,在-20℃时,竹柳枝条的相对电导率值己经达到 50%以上,说明竹柳枝条的半致死温度要低于-20℃;红叶杨枝条的相对电导率在-15℃时相对电导率已达到 55.83%,可判断其枝条的半致死温度要低于-20℃;而竹柳和红叶杨的根系在-10℃时的相对电导率达到了 49.83%和 53.93%,能够判断竹柳根系的半致死温度略高于-10℃,而红叶杨根系的半致死温度要低于-10℃。在-10℃的低温处理条件下,红叶杨和竹柳的枝条以及根系的相对电导率虽逐渐增加,但变化幅度存在差异。如图所示,红叶杨枝条的相对电导率变化幅度较大,较对照组增幅达到了 65.89%,而红叶杨根系的相对电导率较对照组增幅为 10.39%,竹柳枝条的相对电导率较对照组增幅达到了 53.23%,竹柳根系的相对电导率较对照组增幅为 21.95%.
当处理温度在-15℃时,竹柳枝条的相对电导率增加幅度最大,较-10℃处理增加了 48.54%,而红叶杨根系的相对电导率增加幅度最小,较-10℃处理增加了 9.16%,相对而言红叶杨枝条和竹柳根系的增幅较为相似,较-10℃处理分别增加了 17.46%和18.12%;当处理温度在-20℃时,红叶杨根系的相对电导率增加幅度最大,较-15℃的低温处理增加了 25.03%,红叶杨枝条、竹柳枝条以及竹柳根系的相对电导率增加幅度基本相似,较-15℃的低温处理增加幅度分别为 14%、16.78%和 15.85%.当处理温度达到-30℃时,红叶杨和竹柳的枝条以及根系的相对电导率均达到最大值,比对照处理增加幅度分别为 202.89%、67.75%、257.25%、110.86%,其中竹柳枝条的相对电导率增幅最大,红叶杨根系的相对电导率增幅最小。在-25℃--30℃的低温处理过程中,红叶杨和竹柳枝条的相对电导率增幅较大且具有相似性,较-25℃的低温处理,增长幅度分别为 17.93%和 18.43%;相对而言红叶杨和竹柳根系的相对电导率增幅较小且具有相似性,较-25℃低温处理增幅为 2.56%和 5.55%.
焦奎宝利用电导法对葡萄根系的抗寒性研究时发现:随温度的逐渐降低,各种试验材料的相对电导率出现呈“S”状逐渐升高的规律[112].娄晓鸣,吕文涛(2012)等对 10 种景天属植物进行抗寒性研究时指出:随处理温度的逐渐降低,抗寒性较低的景天属植物其相对电导率值较高,并且试验结果与露天试验的结果相符[102].
对红叶杨和竹柳的枝条以及根系在不同低温处理下的相对电导率数据进行方差分析(如表5、表6所示)。根据各供试材料的方差分析数据显示,四组试验材料的相对电导率存在显着性差异。其中,竹柳枝条和红叶杨根系的相对电导率平均数值差异较大,与红叶杨枝条以及竹柳根系相比达到了差异极显着水平。
经过大量研究证实,不同试验数据中的相对电导率数值大小能够相应地反映品种间抗寒能力的大小,即:在同一温度处理条件下,试验数据中的相对电导率数值越小,反映其对低温的抗性越强;试验数据中的相对电导率数值越大,反映其对低温的抗性越弱。
因此,在比较试验材料的抗寒性能力强弱时,可根据试验数据进行系统分析。在本次试验研究中,通过对试验材料的电导率数据比较分析可得出抗寒性强弱:竹柳枝条>红叶杨枝条;竹柳根系>红叶杨根系(即竹柳>红叶杨)。根据试验中相对电导率的变化趋势总结出:即对于两种植物而言,竹柳抗寒性强于红叶杨抗寒性;对于同种植物而言,枝条的抗寒性强于根系的抗寒性。
如图 6 所示,随着处理温度的逐渐降低,供试材料的丙二醛(MDA)含量的变化趋势基本相似,均呈现逐步升高的趋势,其中红叶杨根系的丙二醛(MDA)含量分别在 CK、-10℃、-20℃、-25℃、-30℃等低温处理条件下为最高,含量分别为1.39mol/g、2.13mol/g、2.67?mol/g、2.78mol/g、2.93mol/g,较对照组增幅分别达到了 53.23%、92.08%、100.00%、和 110.79%;其中在 CK--10℃的低温胁迫处理过程中增加幅度较其他组温度处理最为明显。在-15℃的低温处理过程中,红叶杨根系的丙二醛(MDA)含量首次低于竹柳根系,并且在-15℃--20℃的低温胁迫处理条件下,丙二醛(MDA)含量增加幅度趋于缓慢,较-15℃的低温处理增加幅度为 8.53%.
而竹柳枝条的丙二醛(MDA)含量分别在 CK、-10℃、-15℃、-20℃、-25℃、-30℃等温度处理下始终为最低,含量分别为 1.27mol/g,1.58mol/g,2.16mol/g,2.57mol/g,2.63mol/g,2.74mol/g,较对照组增幅分别达到了 24.40%,70.07%,102.36%和 115.74%;其中在-10℃--15℃的低温处理过程中涨幅最为明显,较-10℃的低温处理涨幅达到了 36.7%;在-20℃--25℃的低温胁迫处理过程中丙二醛(MDA)含量增幅最为缓慢,较-20℃的低温处理丙二醛(MDA)含量增幅为 2.33%.
竹柳根系在-10℃--15℃低温处理过程中丙二醛(MDA)含量增幅较为明显,较-10℃的低温胁迫处理丙二醛(MDA)含量增加幅度为 46.24%;而在-15℃--30℃丙二醛(MDA)含量增幅趋于平缓,在-15℃--20℃的低温胁迫过程中,竹柳根系的丙二醛(MDA)含量从 2.53?mol/g 增加到 2.66?mol/g,增幅为 5.13%,并且在-15℃的低温处理其丙二醛(MDA)含量为最高。在-20℃--25℃的低温胁迫处理过程中丙二醛(MDA)含量的增加幅度在整个低温处理过程中增幅为最低,较-20℃的低温处理增幅仅为 0.75%.
红叶杨枝条在整个低温处理过程中其丙二醛(MDA)含量在-10℃--15℃增幅较为明显为 53.23%,在-15℃--30℃的低温处理过程中丙二醛(MDA)含量增幅较为缓慢,其中当低温处理在-20℃--25℃的低温处理阶段增幅最为缓慢,较-20℃的低温处理增幅仅为 1.14%.
红叶杨和竹柳的枝条及根系在低温胁迫过程始末丙二醛(MDA)的含量增幅分别为 112.87%、110.79%和 115.74%、114.17%.
苏向辉,秦伟等(2012)对 4 种李属树种抗寒性分析时发现:随着处理温度的逐渐下降,各试验品种丙二醛(MDA)含量出现逐渐增加的趋势,经过对试验数据系统分析能够得出丙二醛(MDA)含量增加量与增加幅度与各品种植物抗寒性的关系[104].
3.2 低温胁迫对红叶杨和竹柳丙二醛(MDA)含量的影响。
如图 6 所示,随着处理温度的逐渐降低,供试材料的丙二醛(MDA)含量的变化趋势基本相似,均呈现逐步升高的趋势,其中红叶杨根系的丙二醛(MDA)含量分别在 CK、-10℃、-20℃、-25℃、-30℃等低温处理条件下为最高,含量分别为1.39mol/g、2.13mol/g、2.67mol/g、2.78mol/g、2.93mol/g,较对照组增幅分别达到了 53.23%、92.08%、100.00%、和 110.79%;其中在 CK--10℃的低温胁迫处理过程中增加幅度较其他组温度处理最为明显。在-15℃的低温处理过程中,红叶杨根系的丙二醛(MDA)含量首次低于竹柳根系,并且在-15℃--20℃的低温胁迫处理条件下,丙二醛(MDA)含量增加幅度趋于缓慢,较-15℃的低温处理增加幅度为 8.53%.
而竹柳枝条的丙二醛(MDA)含量分别在 CK、-10℃、-15℃、-20℃、-25℃、-30℃等温度处理下始终为最低,含量分别为 1.27?mol/g,1.58?mol/g,2.16?mol/g,2.57?mol/g,2.63?mol/g,2.74?mol/g,较对照组增幅分别达到了 24.40%,70.07%,102.36%和 115.74%;其中在-10℃--15℃的低温处理过程中涨幅最为明显,较-10℃的低温处理涨幅达到了 36.7%;在-20℃--25℃的低温胁迫处理过程中丙二醛(MDA)含量增幅最为缓慢,较-20℃的低温处理丙二醛(MDA)含量增幅为 2.33%.
竹柳根系在-10℃--15℃低温处理过程中丙二醛(MDA)含量增幅较为明显,较-10℃的低温胁迫处理丙二醛(MDA)含量增加幅度为 46.24%;而在-15℃--30℃丙二醛(MDA)含量增幅趋于平缓,在-15℃--20℃的低温胁迫过程中,竹柳根系的丙二醛(MDA)含量从 2.53?mol/g 增加到 2.66?mol/g,增幅为 5.13%,并且在-15℃的低温处理其丙二醛(MDA)含量为最高。在-20℃--25℃的低温胁迫处理过程中丙二醛(MDA)含量的增加幅度在整个低温处理过程中增幅为最低,较-20℃的低温处理增幅仅为 0.75%.
红叶杨枝条在整个低温处理过程中其丙二醛(MDA)含量在-10℃--15℃增幅较为明显为 53.23%,在-15℃--30℃的低温处理过程中丙二醛(MDA)含量增幅较为缓慢,其中当低温处理在-20℃--25℃的低温处理阶段增幅最为缓慢,较-20℃的低温处理增幅仅为 1.14%.
红叶杨和竹柳的枝条及根系在低温胁迫过程始末丙二醛(MDA)的含量增幅分别为 112.87%、110.79%和 115.74%、114.17%.
苏向辉,秦伟等(2012)对 4 种李属树种抗寒性分析时发现:随着处理温度的逐渐下降,各试验品种丙二醛(MDA)含量出现逐渐增加的趋势,经过对试验数据系统分析能够得出丙二醛(MDA)含量增加量与增加幅度与各品种植物抗寒性的关系[104].
以试验材料为不同处理,以在不同温度胁迫下的丙二醛(MDA)含量为试验重复进行单因素方差分析。根据数据丙二醛(MDA)含量平均值排序分析得出:红叶杨根系>竹柳根系>红叶杨枝条>竹柳枝条。对所有试验材料的丙二醛(MDA)含量进行方差和显着性分析得出(如表 7、表 8 所示):红叶杨枝条以及根系的丙二醛(MDA)含量高于竹柳枝条以及根系的丙二醛(MDA)含量,并且红叶杨根系和竹柳枝条丙二醛(MDA)含量达到了差异显着水平。根据数据中丙二醛(MDA)含量分析得出:红叶杨根系中的丙二醛(MDA)含量最高,表明其抗寒性最差;竹柳枝条的丙二醛(MDA)含量最少,说明其具有较强的抗寒性。根据各试验材料中丙二醛(MDA)含量分析比较可得出抗寒性排序:竹柳枝条>红叶杨枝条;竹柳根系>红叶杨根系(即竹柳>红叶杨)根据试验中丙二醛(MDA)含量的变化规律得出结论:即对于两种植物而言,竹柳抗寒性强于红叶杨抗寒性;对于同种植物而言,枝条抗寒性大于根系的抗寒性。





