3.6 低温胁迫对红叶杨和竹柳过氧化物酶(POD)含量的影响。
如图 10 所示,供试材料在低温胁迫处理过程之中,POD 酶活性变化均表现出先上升后下降的变化规律 .随着处理温度的逐渐下降,POD 酶活性初始阶段逐渐升高,由于供试材料抗寒能力不同,其 POD 酶活性的峰值出现的时间也存在差异。
竹柳的枝条以及根系在 CK--15℃时,POD 酶活性持续升高,并且在-15℃时达到最大值;其枝条在这段温度处理过程中 POD 酶活性增幅非常明显,在-10℃和-15℃时,POD 酶活性分别为 14.23U/g·FW 和 18.21U/g·FW,较对照组增幅分别为 32.86%和 70.02%;其根系在-10℃和-15℃时,POD 酶活性分别为 13.53U/g·FW 和18.37U/g·FW,增幅分别为 54.45%和 109.70%.
竹柳枝条以及根系在 CK--10℃的低温处理过程中 POD 酶活性变化规律相似,均呈现逐渐增加的规律,但增幅存在差异。竹柳枝条的 POD 酶活性由 11.71U/g·FW增加到 14.23U/g·FW,增幅为 21.52%;而竹柳根系的 POD 酶活性由 8.76U/g·FW 增加到 13.53U/g·FW,增幅为 54.45%.在-10℃--15℃的低温处理过程中,竹柳枝条的POD 酶活性由 14.23U/g·FW 增加到 18.21U/g·FW,增幅为 27.96%;而竹柳根系的 POD酶活性由 13.53U/g·FW 增加到 18.37U/g·FW,增幅为 35.77%.因此,在 CK--15℃的低温处理过程中,竹柳枝条以及根系的 POD 酶活性始终保持上升的变化规律,且根系的增加幅度大于枝条的增加幅度。
竹柳枝条以及根系在-15℃时其 POD 酶活性出现顶峰,从-20℃到-30℃区间,其POD 酶活性开始出现逐渐降低的趋势,说明其细胞膜保护酶系统可能在低温胁迫过程中受到了伤害,导致 POD 酶活性降低。
在-15℃--20℃的低温处理过程中,竹柳枝条的 POD 酶活性由 18.21U/g·FW 降低到 15.83U/g·FW,降幅为 13.06%;而竹柳根系的 POD 酶活性由 18.37U/g·FW 降低到 14.55U/g·FW,降幅为 20.79%,即竹柳枝条的 POD 酶活性降幅小于竹柳根系,说明在相同低温处理条件下,竹柳枝条的 POD 酶活性失活程度小于竹柳根系,即-15℃--20℃的低温处理对竹柳根系 POD 酶活性的影响大于竹柳枝条。
在-20℃--25℃的低温处理过程中,竹柳枝条的 POD 酶活性由 15.83U/g·FW 降低到 14.33U/g·FW,降幅为 9.47%;而竹柳根系的 POD 酶活性由 14.55U/g·FW 降低到 14.36U/g·FW,降幅为 1.3%,即竹柳枝条的 POD 酶活性降幅大于竹柳根系。因此,在-20℃--25℃处理过程中,低温对竹柳枝条 POD 酶活性的损伤强于竹柳根系,而酶活性逐渐受到抑制,进而对枝条的影响程度大于根系。
在-25℃--30℃的低温处理过程中,竹柳枝条的 POD 酶活性由 14.33U/g·FW 降低到 12.98U/g·FW,降幅为 9.42%;而竹柳根系的 POD 酶活性由 14.36U/g·FW 降低到 11.22U/g·FW,降幅为 21.86%,即竹柳枝条的 POD 酶活性降幅小于竹柳根系。
红叶杨的枝条以及根系在 CK--20℃时,POD 酶活性持续升高,并且在-20℃时达到最大值,之后在-20℃--30℃的低温处理过程中,POD 酶活性逐渐减弱。
在 CK--10℃的低温处理过程中,红叶杨枝条的 POD 酶活性由 9.23U/g·FW 升高到 13.67 U/g·FW,增幅为 48.1%;红叶杨根系的 POD 酶活性由 8.45U/g·FW 增加到13.03U/g·FW,增幅为 54.2%,即红叶杨枝条的 POD 酶活性增幅小于红叶杨根系。
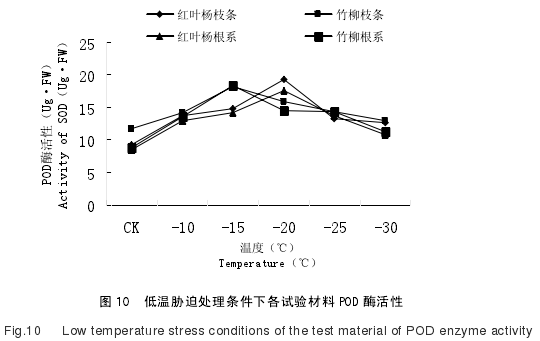
在-10℃--15℃的低温处理过程中,红叶杨枝条的 POD 酶活性由 13.67U/g·FW 升高到 14.86 U/g·FW,增幅为 8.70%;红叶杨根系的 POD 酶活性由 13.03U/g·FW 增加到 14.21U/g·FW,增幅为 9.05%,即红叶杨枝条的 POD 酶活性增幅小于红叶杨根系。
在-15℃--20℃的低温处理过程中,红叶杨枝条的 POD 酶活性由 14.86U/g·FW 升高到 19.23 U/g·FW,增幅为 29.40%;红叶杨根系的 POD 酶活性由 14.21U/g·FW 增加到 17.66U/g·FW,增幅为 24.27%,即红叶杨枝条的 POD 酶活性增幅大于红叶杨根系。
在-20℃--25℃的低温处理过程中,红叶杨枝条的 POD 酶活性由 19.23U/g·FW 降低到 13.26 U/g·FW,降幅为 31.04%;红叶杨根系的 POD 酶活性由 17.66U/g·FW 降低到 13.76U/g·FW,降幅为 22.08%,即红叶杨枝条的 POD 酶活性降幅大于红叶杨根系。
在-25℃--30℃的低温处理过程中,红叶杨枝条的 POD 酶活性由 13.26U/g·FW 降低到 12.58U/g·FW,降幅为 5.12%;红叶杨根系的 POD 酶活性由 13.76U/g·FW 降低到 10.73U/g·FW,降幅为 22.02%,即红叶杨枝条的 POD 酶活性降幅小于红叶杨根系。
如表15、16所示,以试验材料为不同处理,以在不同温度胁迫下的 POD 酶活性为试验重复进行单因素方差分析。根据数据 POD 酶活性平均值排序分析得出:竹柳枝条>红叶杨枝条>竹柳根系>红叶杨根系。对所有试验材料的 POD 酶活性进行方差和显着性分析得出,竹柳枝条以及根系的 POD 酶活性高于红叶杨枝条以及根系的POD 酶活性,竹柳的枝条与红叶杨根系达到了差异极显着水平。根据低温胁迫过程中所有试验材料中的 POD 酶活性数据进行系统分析得出: 红叶杨根系中的 POD 酶活性最低,表明其抗寒性最差;竹柳枝条的 POD 酶活性最高,表明其具有较强的抗寒性。根据各试验材料中 POD 酶活性分析比较可得出抗寒性排序:竹柳枝条>红叶杨枝条;竹柳根系>红叶杨根系根据试验中 POD 酶活性的变化规律得出结论:即对于两种植物而言,竹柳抗寒性强于红叶杨抗寒性;对于同种植物而言,枝条抗寒性大于根系的抗寒性。
3.7 红叶杨和竹柳的抗寒性隶属函数综合评价。
植物的抗寒性受生理生化特征综合影响, 单一的抗寒指标难以对植物的抗寒能力进行综合评价。因此,利用隶属函数分析法可以在多项指标综合测定的基础上对植物的抗寒性进行综合评价[106].
对红叶杨和竹柳的枝条以及根系的相对电导率、丙二醛(MDA)含量、超氧化物歧化酶(SOD)活性、过氧化物酶(POD)活性、可溶性糖含量、可溶性蛋白质含量等 6 项试验数据用模糊数学隶属函数法进行分析。并根据各指标和的平均值进行抗寒性综合评定,对其抗寒性进行了排序。通过比较不同试验材料的生理指标隶属函数平均值对红叶杨以及竹柳(枝条以及根系)的抗寒性进行综合分析,方法如下所示:当试验中某指标与抗寒性呈正相关时,则公式为:N正相关= ( N-N最小) /( N最大- N最小)当试验某指标与抗寒性呈负相关时,则公式为:N负相关=1-(N- N最小)/( N最大- N最小)N 为某一测定数据的数值;N最小为相同测定数据中的最小值;N最大为相同测定数据中的最大值。对红叶杨和竹柳的各统计数据进行累计并计算平均值。通过按照大小顺序对红叶杨和竹柳(枝条以及根系)的隶属函数平均值进行排序比较,进而总结出各试验材料的抗寒性强弱[107].
如表17所示,根据数据得出抗寒性综合排名:竹柳枝条>红叶杨枝条>竹柳根系>红叶杨根系(即:竹柳的抗寒性强于红叶杨的抗寒性)
3.8 红叶杨和竹柳的测定指标与温度之间的相关性分析。
对红叶杨枝条各生理指标与温度进行相关性分析结果表明(如表18所示):温度与相对电导率、丙二醛(MDA)含量、可溶性糖含量、可溶性蛋白质含量、SOD 酶活性、POD 酶活性均呈负相关,相关系数分别为-0.989、-0.946、-0.962、-0.351、-0.034和-0.195,其中与相对电导率、丙二醛(MDA)、可溶性糖呈显着性负相关。相对电导率分别与其他生理指标呈正相关,与丙二醛(MDA)含量和可溶性糖含量呈显着性正相关,相关系数为0.954和0.954.丙二醛(MDA)与可溶性糖呈显着性正相关,相关系数为0.938.SOD 与 POD 呈显着性正相关,相关系数为0.939.
对红叶杨根系各生理指标与温度进行相关性分析结果表明(如表19所示):温度与相对电导率、丙二醛(MDA)含量、可溶性糖含量、可溶性蛋白质含量、POD 活性均呈负相关,相关系数分别为-0.973、-0.914、-0.969、-0.346、和-0.179,与 SOD活性呈正相关,相关系数为0.196.相对电导率与 SOD 酶活性呈负相关,与可溶性糖含量呈显着性正相关,相关系数分别为-0.205和0.983.可溶性糖与 MDA 呈显着性正相关,相关系数为0.917.POD 酶与 SOD 酶呈显着性正相关,相关系数为0.899.
对竹柳枝条各生理指标与温度进行相关性分析结果表明(如表20所示):温度与相对电导率、丙二醛(MDA)含量、可溶性糖含量、可溶性蛋白质含量、SOD 活性、POD 活性均呈负相关,相关系数分别为-0.993、-0.978、-0.946、-0.889、-0.0406和-0.325.
相对电导率与 MDA 含量和可溶性糖含量呈显着性正相关,相关系数分别为0.989和0.926.丙二醛(MDA)与可溶性糖和可溶性蛋白质呈显着性正相关,相关系数为0.942和0.917.可溶性蛋白质和可溶性糖呈显着性正相关,相关系数为0.981.
对竹柳根系各生理指标与温度进行相关性分析结果表明(如表21所示):温度与相对电导率、丙二醛(MDA)含量、可溶性糖含量、可溶性蛋白质含量、POD 活性均呈负相关,相关系数分别为-0.996、-0.931、-0.964、-0.845和-0.283,与 SOD 活性呈正相关,相关系数为0.163.相对电导率与丙二醛(MDA)含量和可溶性糖含量呈显着性正相关,相关系数分别为0.941和0.974.丙二醛(MDA)与可溶性糖呈显着性正相关,相关系数为0.955.可溶性糖和可溶性蛋白质呈显着性正相关,相关系数为0.941.
3.9 红叶杨和竹柳枝条恢复生长分析。
恢复生长法具有可靠且易于比较的特点,并且可以作为其他研究性方法的对照,因此在对红叶杨和竹柳的抗寒性研究中使用。恢复生长法是将经不同低温处理过的离体枝条放入具一定条件的组培室内进行水培,通过观察枝条的萌芽情况,进而对数据进行统计分析,最终确定试验材料的致死临界温度。
根据红叶杨和竹柳枝条的萌芽率可得出(如表22所示),从对照组开始至-30℃整个低温胁迫过程中,两品种的枝条萌芽率均呈现不同程度递减趋势。红叶杨枝条从对照组开始到-20℃枝条的萌芽率从55.7%递减到0,在 CK--10℃过程中,萌芽率较对照组降低28.9%.当胁迫温度处理在-15℃时,萌芽率仅为6.5%,较对照组降低88.33%,说明低温胁迫已达到红叶杨的生理极限,部分生理组织受到损伤,枝条的正常萌芽状态受到外界环境刺激而逐渐丧失萌芽能力。当温度处理达到-20℃时,红叶杨枝条已不具备萌芽能力。
竹柳枝条从对照组开始到-25℃枝条的萌芽率从 63.5%递减到 0,在 CK--10℃过程中,萌芽率较对照组降低 32.59%.当胁迫温度处理在-15℃时,萌芽率为 18.2%,较对照组降低 77.33%,说明低温胁迫对竹柳的生理组织造成严重损伤,导致萌芽率骤降。当处理温度达到-20℃时,竹柳的萌芽率仅为 3.5%,说明受低温胁迫的影响,竹柳枝条的正常萌芽状态已达到峰值。当温度处理达到-25℃时,红叶杨枝条已不具备萌芽能力。
根据枝条的恢复生长试验数据可得出:当外界温度低于-15℃时,红叶杨的正常萌芽状态受到影响,外界低温会对红叶杨枝条的正常萌芽产生抑制作用。当外界温度保持在-20℃及以上时,竹柳枝条会保持正常的萌芽状态。因此,通过枝条的萌芽率进一步总结出竹柳的抗寒性强于红叶杨的抗寒性。





