
引言
坐落于斯匹次卑尔根群岛上的新奥尔松地区,受大西洋暖流影响,是北极最温暖的地区之一。该地区气候温和,即使在冬季期间,气温也会达到 0℃以上。有监测数据显示:新奥尔松地区气温较其他北极 高 纬 地 区 增 温 更 快,平 均 每 10 年 增 加0. 68℃。它已成为国际上研究北极气候变化的一个常规监测点。
王湾位于斯匹次卑尔根群岛西海岸,是一个半开放性的海湾。它受到了北大西洋暖水团、北极冷水团,及陆地冰川融水的交互影响。夏季,冰川融水伴随着大量的泥沙等颗粒物注入到湾内,从而降低了该海域的真光层深度。冰川融水的注入在一定范围内(距离冰川前沿 45 km)改变了表层海水(30 m 以浅) 的盐度分布。另外,大西洋暖流输送的高温、高盐( >34. 7)海水也对王湾产生了重要影响。Wang 等认为,大西洋暖流在很大程度上影响了湾外40 m 以浅的表层海水的理化特性,从而在很大程度上影响了浮游植物的生长以及该地区浮游生物的群落结构。
光照是影响浮游植物生长的一个关键性因子,而北极浮游植物对光的适应性十分特殊。现有研究表明:某些浮游植物类群在北极寒冷、黑暗的冬季具备生存下来的能力。近几年,国际上对环境变化中的北极浮游植物已做了相关研究。海水盐度的下降和泥沙的注入是影响北极近岸表层水微型浮游生物群落组成和多样性的决定性因素。Li 等认为,随着北冰洋海水盐度的降低,粒径 <2 μm 的微微型浮游植物可能会大量繁殖,并占据优势地位。
如果这些变化持续下去,这些微微型的浮游生物将会加速“微型生物食物环”的进程,从而改变北冰洋食物链的结构。国内陆续有研究人员对北冰洋海域和中心区进行了浮游植物类群改变和环境变化的相关性分析。然而,有关浮游植物对北极环境及其变化的适应性研究很少,且没有王湾相关的记录。
综上所述,本研究于现场实验的基础上评估王湾浮游植物对光照和盐度变化的适应性,藉此弄清两个问题:(1)浮游植物是否能够适应黑暗环境;(2)冰川融水的注入,对浮游植物生长有何影响。
1 材料和方法
1. 1 环境数据和样品采集
2006 年 8 月初,在斯匹次卑尔根群岛新奥尔松地区的王湾海域确定了由 5 个站位组成的一条断面(图 1)。利用经校正的 SEB 19 plus CTD 测定了站位的盐度、光合有效辐射(photosynthetically activeradiation,PAR)和叶绿素 a 浓度剖面。用 2. 5 L 的Niskin 采样瓶采集 5 m 水层的水样。水样运回新奥尔松海洋实验室,并立即做实验处理。用于盐度实验的冰川融水采集于新奥尔松机场附近的融水溪流;为了去除大量的泥沙等颗粒物,将采集到的融水静置于实验室外的平台 1 天时间。开始实验前,各实验分别取 500 mL 水样用于叶绿素 a 浓度本底的测定。【图略】
1. 2 暗适应实验
1. 2. 1 培养周期 1 天将水样转移到 5 个容积为 500 mL、经稀盐酸浸泡晾干的 Nelgene 培养瓶中,其中 3 个培养瓶用铝箔包裹以保持黑暗的培养环境,另外 2 个暴露于自然光照条件下。培养实验在码头附近的一个“实验区”进行,培养 1 天后将每个培养瓶摇匀,各取200 mL用于叶绿素 a 浓度的测定。
1. 2. 2 培养周期 10 天实验采用两个平行样。在第一阶段,将浮游植物样品瓶放置于海洋实验室的培养槽中,并保持完全的黑暗环境。为了保持培养环境的温度和实际环境中的温度基本一致,培养中循环水为实验室内的采水系统于海湾中 70 m 深采集的连续供应海水。培养瓶每天 1 次进行摇匀。培养 7 天后,混合均匀并取100 mL 水样用于叶绿素 a 浓度的测定。而后将培养实验移到码头附近的实验区,继续在自然光照下培养 3 天;培养 1 天和培养结束时,分别取100 mL 和200 mL 样品用于叶绿素 a 浓度的测定。
1. 3 盐度实验
实验水样采集于 S2 站和 S4 站。针对每个站位的样品,各设置 6 个不同盐度梯度,海水和冰川融水的混合体积比分别为 100∶ 0、90∶ 10、80∶ 20、60∶ 40、40∶ 60 和 20 ∶ 80。每个培养实验设置 3 个平行样。在码头附近实验区培养 1 天后,每个培养瓶各取200 mL 样品用于叶绿素 a 浓度的测定。
1. 4 叶绿素 a 浓度的测定
水样用 GF/F 滤膜(Ф 47 mm,孔宽 0. 7 μm)过滤,收集滤膜并在低温环境中(0—4℃)用 90% 的丙酮溶液萃取 24 h,按 Parsons 等的方法,利用唐纳荧光仪(TURNER10-AU 型)测定叶绿素 a 浓度。
1. 5 浮游植物生长率的计算
浮游植物生长速率的计算公式如下:【1】
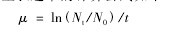
其中,μ是指浮游植物生长速率,Nt是培养后浮游植物生物量(叶绿素 a 浓度代替),N0是开始培养之前浮游植物生物量(叶绿素 a 浓度代替),t 是培养时间(单位:d)。
2 结果
2. 1 研究区域的环境特征
图 2 所示为采样站位上层海水(50 m 以浅)盐度、光合有效辐射(PAR)、叶绿素 a 浓度的分布情况。受到冰川融水的影响,表层盐度较低,且随着水深的增加而升高,并且出现了明显的盐度跃层。在5 m 层,盐度从湾外到湾内呈现降低的趋势,但最低值出现在 S2 站(31. 43)。S1 站、S2 站,PAR 随水深的增加而快速减弱,水深 10 m 处的 PAR 值低于表层的 10%。【图.略】
湾内海水的浑浊度较湾外高。真光层深度从湾外到湾内呈降低的趋势,其深度在 25—30 m 之间。除了在湾外 S1 站,叶绿素 a 最大值出现在水深20 m处,其他站位叶绿素 a 浓度的最高值均出现在 10—15 m 的水层。
2. 2 暗适应实验
在光照和黑暗条件下培养 1 天后,叶绿素 a 平均浓度(3 个平行样)没有出现明显的差异,说明浮游植物群落在黑暗条件下仍可以生存。采集于湾外S1 站和 S2 站的水样,光照条件下培养的浮游植物生长速率分别为 -0. 04 和 0. 03,而在黑暗条件下,其生长速率分别为 -0. 31 和 -0. 1(表 1),可见,黑暗条件下培养对浮游植物生长的影响更为明显。湾内采集的浮游植物群落在黑暗和光照条件下培养,其生长速率没有明显差异。图 3 显示了不同站位采集的浮游植物群落在光照和黑暗环境下培养 1 天后的叶绿素 a 浓度之比,从湾外到湾内,比率呈现明显下降的趋势,表明黑暗环境对湾外浮游植物群落生长的影响更为明显。在黑暗环境下经过 7 天的培养后,叶绿素 a 浓度没有出现较大的变化。将培养实验移到自然光照条件下培养 1 天后,叶绿素 a 浓度出现明显下降,而后又开始快速增长。在 S2 站,在整个培养周期内,浮游植物群落生长速率由 0. 029 下降到 -0. 126,培养结束后,生长速率又增加到 0. 088 5。然而,在 S4站,生长速率由 -0. 003 下降到 -0.042,在培养结束时增加到 0. 163。【表1 .图3】
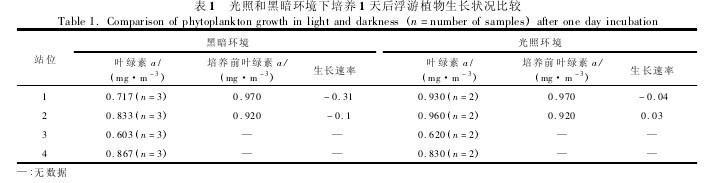
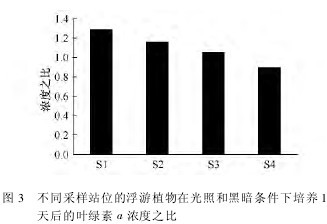
图 4 所示的结果表明,与 S4 站比较,采自于湾外 S2 站的浮游植物表现出更好的对黑暗环境的适应能力。这有可能归因于两个站区的浮游植物群落组成和生物量不同。Wang 等研究指出,湾内和湾外表层海水存在不同的环境水平和生物类群组成,微型的异养鞭毛虫生物量在湾外表层水中较高。【图4】
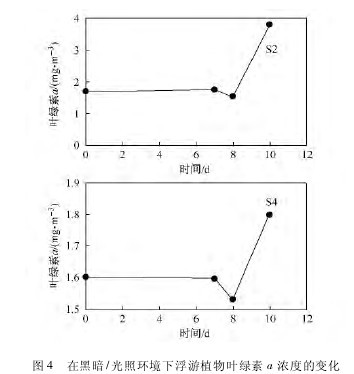
硅藻对湾外浮游植物生物量的贡献最大,然而在湾内,自养腰鞭毛藻丰度和多样性较高。2006 年8 月,湾内的浮游植物有可能受到大量冰川融水注入的影响,导致真光层深度变浅(图 2),从而抑制了其生长。大量的泥沙等颗粒物的注入导致湾内的浮游植物受到了弱光照的影响,这可能导致湾内的浮 游 植 物 生 物 量 (1. 6 mg·m- 3) 较 湾 外 的 低(1. 8 mg·m- 3)。
另外,在黑暗环境中经过 7 天培养后,当将培养暴露于自然光下后,浮游植物生物量出现急剧的下降,这表明强光照对浮游植物群落中部分“暗适应”能力较强的生物类群的生长产生了抑制作用。
2. 3 盐度实验
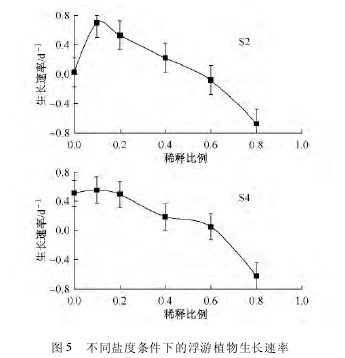
图 5 显示在模拟海水温度和自然光照下,浮游植物群落在不同的盐度环境中,其生长速率随盐度的变化曲线。当稀释比例(冰川融水和海水的混合体积比)为 10∶ 90 时,浮游植物群落生长速率最高,当稀释比例增加或降低而使盐度值减少或增加时,浮游植物生长速率随之减低。稀释比例为 0 时,浮游植物生长速率较稀释比例为 20∶ 80 的高;稀释比例为 80∶ 20 时,所观察到的生长速率最低。在 S2站,当稀释比例达 60∶ 40 时,浮游植物群落生长速率和未作稀释的培养样中的生长速率接近;而在 S4站,出现相似情况时,只需要稀释比例达 20∶ 80 即可。这也反映出湾内的浮游植物群落在盐度实验前就已受到冰川融水更为显着的影响。【图5】
3 讨论
3. 1 浮游植物对黑暗环境的适应能力
有报道指出,高纬海域的浮游植物对暗环境具有一定适应能力。本研究的结论也佐证了前人的研究,说明高纬海域浮游植物群落能在黑暗环境中生存。利用荧光显微分析对冬季(极夜)北冰洋中心区调查研究显示:粒径在 2 μm 左右的浮游植物类群大量存在,并且在之后的春季浮冰冰下水华浮游植物群落中占据较大的比重。连续的季节性观测显示:浮游植物某些类群能在冬季极夜条件下生存,能够在较低浓度下保持自身色素的完整性,以便在晚冬或早春当太阳光出现时,浮游植物群落能够生长、繁殖。其中,混合营养型浮游植物具备两种生活方式:既能进行光合自养,又能靠异养方式从外界获取能量。这可能是该类浮游植物能在低光照强度的高纬度海域生存的原因之一。混合营养型在极地海洋或淡水环境中普遍存在,多数研究认为该类群在极区非常重要。定鞭藻类或者其他冰下微藻具备叶绿体,可能都是属于混合营养型浮游植物,其营养方式可能包括“渗透营养型”和“吞噬营养型”。能够适应低光照强度的嗜冷性浮游植物存在于极地海洋中,观测指出,当将嗜冷性浮游植物从低光照强度转到高光照强度的环境中后,会导致其光合效率下降。
3. 2 盐度变化对浮游植物生长的影响
Fredersdorf 等指出,一种北冰洋褐藻 Alariaesculenta 能够适应和忍受一定范围内的盐度变化。
研究表明,盐度变化对南北极冰藻的生长及浮游植物群落组成都具有很大程度的影响。在一定程度上,冰藻能适应盐度的降低,然而,北冰洋浮游植物群落能否适应较低的盐度值得进一步研究。在湾内和湾外的不同站位之间,浮游植物群落生长速率不同;我们认为可能是因为不同站位间,浮游植物群落组成有差异,尽管没能够研究该海湾浮游植物群落组成,以及在不同盐度下,培养前后的浮游植物组成变化情况。王湾是一个小海湾,其中一面是开阔海域。在海湾中部形成一海底山脊,并将海湾分成了两部分,其中,湾外主要受到斯匹次卑尔根暖流的影响,而湾内主要受到冰川融水的影响。湾内复杂水团的交汇作用是导致浮游植物群落组成变化的主要因素。硅藻是对浮游植物生物量贡献最大的类群,特别是在湾外海域。然而,自养腰鞭毛藻是湾内浮游植物生物量最主要的贡献者。研究表明:在表层水中,腰鞭毛藻和金藻 Dinobryon balticum占主导地位,而硅藻和定鞭藻 Phaecystis pouchetii 的丰度则会随水深的增加而增加。显微镜和分子生物学技术分析显示:2006 年度夏期间,王湾腰鞭毛藻多样性较高。
如图 5 所示,在较低的稀释比例下,采自 S4 站的浮游植物生长速率较 S2 站的稳定。这反映出受冰川融水的影响,从湾外到湾内的盐度逐渐降低。
和湾内比较,湾外的浮游植物群落对淡水输入所导致的盐度变化更敏感。Wang 等研究指出,整个湾表层水的盐度值较低(31. 5),受冰川融水的影响,湾内的盐度值更低。随着所添加冰川融水量的不同,浮游植物生长速率也会不同,该现象可能由两方面原因导致。
(1) 有利方面 冰川融水的添加减少了浮游动物对浮游植物的摄食压力,从而促进了浮游植物的生长。另外,盐度变化会影响浮游动物的存活率,也将减少浮游动物的摄食压力。同时,不同量融水的添加,改变了各组样品中营养盐含量的水平。融水的添加,使得浮游植物可吸收的营养盐含量增加。然而,本研究的培养时间较短、浮游植物的生长速率也不是很高,营养盐不应成为浮游植物生长的限制因子。
(2) 不利方面 由于培养样品受到融水的稀释,自然光照强度减弱,盐度降低,这对浮游植物的生长起了抑制作用。由本研究的暗实验结果可见,浮游植物群落具有一定的暗环境适应能力。而盐度实验显示,当融水和海水的混合比例增加到 80∶ 20 时,浮游植物的生长速率减小到最低值,这表明盐度是影响浮游植物生长的一个关键因子。大量融水的输入降低了表层海水盐度、温度,融水成为其他微型生物、污染物和有机碳的来源之一,对浮游植物影响过程可能极为复杂。本研究的模拟实验是基于王湾海域夏季受大量冰川融水注入的实际而制定的,若要深入了解冰川融水注入对浮游植物群落的影响,应在群落结构变化等方面开展更为深入的研究。
4 结论
通过模拟实验揭示,采自于王湾的浮游植物群落在暗光条件下仍能够生存,从侧面反映出该浮游植物群落具备一定的暗适应能力,另外,适量的冰川融水所导致的盐度变化能够促进浮游植物的生长,但是更低的盐度则会抑制浮游植物的正常生长,暗示了北极持续变暖所导致冰川融水的大量注入可能会对浮游植物生长不利。由于现场条件的限制,初次模拟实验中,本研究只分析了浮游植物叶绿素 a的变化情况,未来开展实验时,将会着重分析初级生产力、浮游植物组成及其数量的变化,并以此来评估北极环境变化对浮游植物的影响。





