
1 引言
20世纪中叶以来,随着矿物燃料燃烧、化学氮肥的生产和使用、畜牧业的迅猛发展,人类活动向大气中排放的活性氮化合物激增,使得大气氮素沉降也呈迅猛增加的趋势。与此同时,伴随着全球变化趋势的加剧,降水量在全球尺度上也发生了明显变化,Zhang等指出,北半球中纬度地区气候和南半球亚热带地区的降水量明显增加。由于大气氮沉降通过对植物生物量与土壤活性碳动态的直接或间接作用,极大地干预了陆地生态系统碳循环和碳蓄积过程,而降水是干旱、半干旱生态系统重要的水分来源,全球变化导致的降水格局变化则从不同尺度影响植物生理及生态学过程,因此,氮素沉降和降水对生态系统碳蓄积的影响是近年来全球变化研究的热点之一。
构件是植物体上具有生死过程和潜在分生能力的形态学单位,它的体积大小基本可以代表生物量储量。由于受到水分、养分、温度等环境因子的影响,植物将资源分配于各构件时会进行权衡,使生长、防御和繁殖等方面的效益达到最佳。近年来,关于氮沉降和降水变化对植物生物量储量的影响受到普遍关注。有研究表明,在氮缺乏的生态系统中,氮输入的增加能刺激植物生长,提高生产力,但长期输入氮素却使植物生物量减小;在氮饱和的生态系统,氮的输入还会抑制植物的生长,甚至导致生态系统衰退。也有研究表明,短命植物群落的生产力与降水量密切相关。降水量大的年份里处在旺盛生长阶段的短命植物的生物量(风干重)可占到所有植物总生物量58%。但是,大多数研究侧重于群落或种群生物量对氮素和水分的响应,有关水分和氮素添加对植物个体生物量分配格局的影响研究较少。因此,本文着重研究了氮素及水分添加对科尔沁沙地优势植物地上生物量分配的影响,以期为预测未来气候变化背景下干旱及半干旱地区生态系统的演替动态及可持续发展提供科学依据。
2 研究区域自然概况
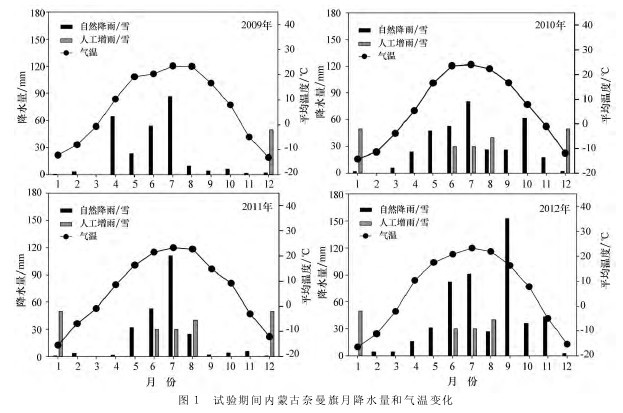
研究区域位于科尔沁沙地中南部奈曼旗境内。地理位置为42°58′N、120°44′E,海拔约360m。年均气温6.5℃,最热月(7月)平均气温23.5 ℃,最冷月(1月)平均气温-13.2℃,全年≥10℃的有效积温3 200~3 400℃,无霜期151d,极端最低气温-29.3℃,夏季无植被覆盖的沙丘表面最高温度可达57.2~60.0 ℃。年 太 阳 辐 射 总 量 为5 200~5 400MJ·m-2,多年平均降水为360mm,主要集中在6—8月,年蒸发量1 935mm,属温带大陆性半干旱气候类型。图1为该地区2009—2012年试验期间月降水量和平均气温变化情况。该地区的地貌类型以固定沙丘、半固定沙丘、流动沙丘和丘间甸子交错分布为主。优势植物以差不嘎蒿(Artemisia halodendron)、小叶锦鸡儿(Cara-gana microphylla)等 灌 木 和 半 灌 木 以 及 狗 尾 草(Setaria viridis)、糙隐子草(Cleistogenes squarro-sa)、胡枝子(Lespedeza davurica)和扁蓿豆(Melis-silus ruthenicus)等一年生和多年生的杂草为主。【图1】
3 研究方法
3.1 样地处理和物种选择
研究样地位于中国生态系统研究网络奈曼沙漠化研究站附近。地貌特征为相对平坦的缓起伏沙质草地,植被盖度65%~80%。根据需要,在研究样地上设置36个5m×5m试验小区,小区之间留1m宽隔离带,在试验小区内布设对照、水分添加(夏季增雨、冬季增雪)、N素添加、同时添加水分和N素等 试 验 处 理,共 形 成6个 处 理,分 别 为 对 照N0W0、冬季增雪N0W1、夏季增雨N0W2、氮素添加N1W0、冬季增雪×氮素添加N1W1和夏季增雨×氮素添加N1W2,每个处理设6个重复。为了增加研究结果的可比性,参考国际上常用的氮素添加水平,采用中等氮沉降速率水平(100kg·hm-2·a-1),试验期间每年7月初,在添加氮的小区内施入尿素(CN2H4O)535.71g。夏季增雨和冬季增雪的量相当于该地区多年平均降水的30%,约100mm降水,夏 季 增 雨 小 区 每 年6、7、8月 人 工 增 加30mm、40mm、30mm的降水。冬季增雪小区每年11月至次年1月期间,自然降雪后收集样地以外的积雪,分2~3次人工撒入相当于100mm降水的雪量。试验于2009年11月开始,2012年9月结束。
根据研究样地物种的优势程度,选择样地中白草(Pennisetum centrasiaticum)、狗尾草、尖头叶藜(Chenopodium acuminatum)和糙隐子草等4种草本植物为调查对象,进行地上生物量分配研究。其中白草和糙隐子草为多年生禾本科植物,狗尾草为一年生禾本科植物,尖头叶藜为一年生藜科草本植物。
3.2 地上生物量测定
地上生物量测定分别于2010和2012年8月植物个体生物量最大时进行。测定时,在每个处理小区内选定2~3株目标植物个体,然后将每个个体齐地面剪取后装入自封袋并标记,带回实验室后将每个个体分解为繁殖体、茎和叶3部分,在烘箱内70℃下烘干至恒重并称重。植物个体地上总生物量以繁殖体干重、茎干重和叶干重的总和表示;地上生物量分配以各构件生物量干重占个体地上总生物量的百分比表示。
3.3 数据分析
用EXCEL及SPSS进行数据分析,采用多因素方差分析比较施氮、增水及氮水互作处理和年份差异对这4种植物地上部分生物量分配的影响,并用单因素方差分析单个物种对不同处理的响应。用多重比较中的最小显着差数法(LSD)或Tamhane’s检验(方差不齐时)比较各处理间差异。
4 结果与分析
4.1 氮素及水分添加对植物地上总生物量及构件生物量的影响
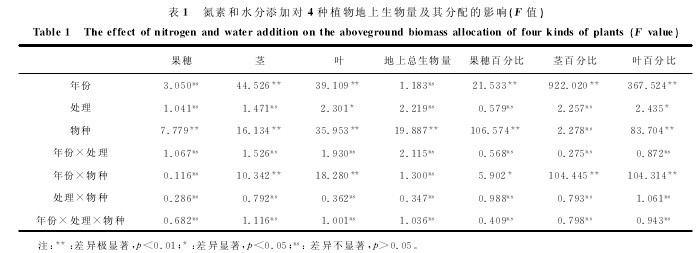
方差分析(表1)显示,添加氮素、冬季增雪、夏季增雨、冬季增雪×氮素添加和夏季增雨×氮素添加等处理下4种植物平均地上总生物量、繁殖体生物量和茎生物量在处理之间无显着差异(p>0.05),但不同处理对叶生物量有显着影响(p<0.05)。由图2可以看出,单纯添加氮素或水分(不论是冬季增雪或夏季增雨)均没有对科尔沁沙地4种优势植物平均地上总生物量及其叶、茎、繁殖体等构件平均生物量产生显着影响(p>0.05),并且冬季增雪×氮素添加处理下,4种植物平均地上总生物量及其构件平均生物量与对照之间亦无显着差异。【表1.图2略】
2010年夏季增雨×氮素添加处理下,4种植物平均地上总生物量及其叶、茎、繁殖体等构件平均生物量显着增加,分别较对照增加了110.84%、93.00%、150.31%和118.38%。但是2012年夏季增雨×氮素处理中平均地上总生物量和构件平均生物量并没有发生显着变化。
分析氮素和水分添加对单个物种地上总生物量及构件生物量的效应,发现各处理对糙隐子草、白草和尖头叶藜地上总生物量及构件生物量无显着影响(p>0.05,图3),但对狗尾草地上及构件生物量具有显着效应。
2010年夏季增雨×氮素添加处理下,狗尾草茎、叶、繁殖体生物量和整株植物的地上总生物量较对照均显着增加 (p<0.05),分别增 加了311.86%、177.32%、229.32%和222.69%。
4.2 氮素及水分添加对地上生物量分配的影响
方差分析显示,添加氮素、冬季增雪、夏季增雨、冬季增雪×氮素添加和夏季增雨×氮素添加等处理下4种植物繁殖体和茎平均生物量分配在处理之间无显着差异(p>0.05),但不同处理对叶平均生物量分配有显着影响(p<0.05,表1)。由图4可以看出,除了在2010年单独增加氮素使叶平均生物量分配显着增加以外(p<0.05),单纯添加氮素或水分(不论是冬季增雪或夏季增雨)均没有对科尔沁沙地4种优势植物平均叶、茎和繁殖体平均生物量分配产生显着影响(p>0.05),并且夏季增雨×氮素添加处理下,4种植物各构件平均生物量分配与对照之间亦无显着差异。
2010年冬季增雪×氮素添加处理下,4种植物叶平均生物量分配较对照显着增加了14.02%。但是2012年冬季增雪×氮素处理中叶平均生物量分配并没有发生显着变化。
分析氮素和水分添加对单个物种各构件生物量分配的效应,发现各处理对糙隐子草、白草和尖头叶藜地上各构件生物量分配无显着影响(p>0.05,图5),但对狗尾草叶生物量分配具有显着效应。
2010年冬季增雪×氮素添加处理下,狗尾草叶生物量分配较对照均显着增加了19.35(p<0.05)。
4.3 年份对4种植物地上生物量及其分配的影响
表1表明,年份对茎、叶平均生物量和茎、叶、繁殖体等构件生物量的分配影响极显着(p<0.01),由图2和图4可知,与2010年相比,2012年的茎平均生 物 量 及 分 配 分 别 极 显 着 增 加 了154.6%和107.5%,而叶平均生物量及分配则极显着减少了67%和46.2%(p<0.01)。
分析不同年份对单个物种各构件生物量分配的效应(图3和图5),与2010年相比,发现除了尖头叶藜以外,其他3种植物的叶和繁殖体生物量及其分配减少极为显着,而茎生物量及其分配均极显着增加(p<0.01)。【图3-5.略】
5 讨 论
以往研究显示,水分和养分是制约半干旱区沙地生态系统植被生产力和受损生态系统植被恢复的重要因子,它们的增加会刺激植物生长,提高生产力,增加生态系统碳蓄积量,这在氮素和水分缺乏的生态系统中具有普遍意义。本试验结果表明,单独添加水分或者氮素对生物量分配没有明显变化,我们认为这可能是由于试验区的土壤为沙土,质地疏松,其持水能力和保肥能力差,地表撒施会使肥料利用率降低,以及由于施水是在每年7—8月,达到年均气温最高点,蒸发量大,从而导致单独增加氮素或水分的对生物量的积累和分配效果不显着。模拟半干旱生态系统的模型表明:如果在水分充足条件下,氮输入能够促进生态系统的生产力;如果水分不足,生产力不会随氮的增加而改变。有研究表明,增加水分更利于植物将更多的生物量分配给叶进行同化产物的积累,但是增水会因为氮素的限制而对叶生物量及其分配的作用有限。在夏季,对荒漠灌木Larrea tridentata进行加水和同时加氮加水这两种处理的结果要比对照和单独加氮处理有更高CO2固定速率、气孔导度和比叶面积,氮素可通过改变氮在叶片中的分配格局影响叶片的光合作用,并通过改变碳水化合物的库源关系和能量消耗水平而调节碳同化物质在体内的分配,同时也解除了氮素对水分限制作用。我们认为这也可能是导致单独增加水分或氮素效果不显着的原因之一。也有研究结果显示,荒漠生态系统通过反硝化作用而损失氮的速率很快,加之由于氮的挥发,使得有效氮的含量降低,从而单独增加氮素的效果不明显。此外,我们考虑只有叶生物量分配对实验处理的响应显着,在一定程度上也说明了资源在不同性状之间分配时存在权衡关系。
试验显示年份差异对生物量的积累和分配影响显着,在2012年地上总生物量和茎生物量的积累、分配均有增加趋势,叶和繁殖体生物量的积累和分配都呈现降低趋势。这可能是由于不同年份植物的生长环境差异很大,如土壤养分、水分,土壤团聚体等环境因子发生改变,引起植物叶性状的改变以适应新的生境条件,地上生物量分配变化也发生改变,将生物量更多的分配给茎可以有效地增加植物对养分和水分的运输,从而更利于叶片进行光合作用以储积更多的生物量。也有研究表明,在干旱半干旱等氮素含量较低地区,水分和养分的增加有可能会因部分原因损失,故在一个短的时期内对促进生物量分配的作用可能不显着,但长期的处理就会产生累积效应使得植物生物量分配显着增加。
此外,2012年7—8月降水量较2010年多,这些都有可能是造成2012年茎生物量的积累和分配显着高于2010年的原因。试验结果表明,不同物种对不同处理的响应差异显着。我们认为,这种差异性既与物种的生活史特征有关,又与干扰的性质有关,这也与以往的研究相同。前人研究也证明种间繁殖输出的差异是环境和遗传因素共同作用的结果。环境因素中,有研究者认为种间这种对干扰响应的差异性是由种间资源限制的不一致性所致,在土壤营养贫瘠的环境中植物对地下营养的竞争是关键的,而对地上光资源竞争则不重要;当群落中土壤营养充足时植物对地下营养的竞争不再十分重要,而对光资源的竞争就显得尤为重要。那些本身茎高叶多,茎叶分配较大、根分配相对较小的物种在对光的竞争中处于优势,并且将会成为群落的优势种,如本实验中的狗尾草。此外,生物量分配的变化与系统发生肯定有着密切关系,但由于所研究物种代表性有限,我们尚无法详尽说明生物量分配的变化与系统发生之间的确切关联。
综上所述,在水分和氮素作为主要限制因子的科尔沁沙地生态系统中,夏季增雨×氮素添加能够解除氮素对水分的限制作用,使植物将所获得的有限资源供于叶片生物量的积累和分配,进行营养生长以增强个体资源获取能力(同化能力)来保证较高的繁殖适合度。此外,不同年份和物种之间本身的差异性也是我们在考虑增水、施氮作用时不可忽略的因素,这种显着的差异有助于我们在全球气候变化的背景下,预测沙地生态系统的稳定程度、更替过程。





