����ժ Ҫ����Ϊ����ȷ������ؽ����������Ժ�������, ��������������, �о�ͭ������ (Microcystis aerugino-sa) �Բ�ͬ����Ũ��Zn2+��Cd2+�ĸ��������Լ�����в�ȵ�������Ӧ���������, ͭ�������Zn2+��Cd2+����һ���ĸ�������, ����Zn2+�ĸ��������Ը���Cd2+����Zn2+����Ũ��Ϊ0. 05 mg/Lʱ, ͭ������ı������������, ��Zn2+����Ũ�ȳ���0. 05 mg/Lʱ, ͭ������������ܵ�����, ��Zn2+��ͭ��������������е�����Ũ�ȴٽ�, ������Ũ�����Ƶ����á�Cd2+��ͭ����������������е�����Ũ�ȴٽ�����, ����Ũ��Ϊ0. 05 mg/L��Cd2+����96 h��, ͭ������������ܵ���������, Cd2+����Ũ��Խ��, ���ͭ��������������������Խ���ԡ�Zn2+��Cd2+����Ũ�ȷֱ�Ϊ0. 20 mg/L��0. 15 mg/Lʱ, ��������24 h��, ͭ���������ø���Ծ���������, ��Zn2+��Cd2+����Ũ�ȷֱ�ﵽ0. 20 mg/L��0. 25 mg/Lʱ, ��������, ��ϸ����ϵͳII (PSII) ������ת��Ч�� (Fv/Fm) ���Ž��͡���Zn2+��Cd2+в����, ͭ������ij��������绯ø (SOD) ���ԡ���������ø (CAT) �����Լ�����ȩ (MDA) �����������仯��
�����ؼ��ʣ���ͭ������; п; ��; ������Ӧ; ��������;
����Abstract����In order to study the tolerance and enrichment characteristics of algae to heavy metals, the enrichment ability and physiological response of Microcystis aeruginosa to different concentrations of Zn2+ and Cd2+ were investigated by using indoor culture methods. The results showed that Microcystis aeruginosa had a certain enrichment effect on Zn2+ and Cd2+ , and the enrichment of Zn2+ in Microcystis aeruginosa was obviously higher than that of Cd2+ . Microcystis aeruginosa grew fastest when the concentration of Zn2+ was 0. 05 mg/L, while the growth was inhibited when the concentration of Zn2+ exceeded 0. 05 mg/L. The low concentrations of Zn2+ could promote the growth of Microcystis aeruginosa, while high concentrations could inhibit its growth, which was called the hormesis effect. However, the growth of Microcystis aeruginosa was not promoted by low concentration of Cd2+ and was obviously inhibited after being treated with 0. 05 mg/L Cd2+ for 96 h. With the increase of Cd2+ concentration, the inhibitory effect of Cd2+ on the growth of Microcystis aeruginosa became more and more obvious. When the concentration of Zn2+ and Cd2+ reached 0. 20 mg/L and 0. 15 mg/L, the esterase activity of Microcystis aeruginosa increased significantly after continuous culture for 24 hours. When the concentration of Zn2+ and Cd2+ reached 0. 20 mg/L and 0. 25 mg/L, respectively, the Fv/Fmvalue was significantly decreased. The activities of superoxide dismutase ( SOD) and catalase ( CAT) and the content of malondialdehyde ( MDA) changed under the stresses of Zn2+ and Cd2+ .
����Keyword����Microcystis aeruginosa; Zn; Cd; physiological response; enrichment ability;
�������Ź�ũҵ�Ŀ��ٷ�չ, �����ؽ�������ˮ����, �ⲻ����Ⱦˮ��, �����������������ڸ���, ��ͨ��ʳ��������, �������÷Ŵ�, ����Σ�����ཡ��[1]��������ˮ����̬ϵͳ�ij���������, ��ϸ���ڼ�ϸ���ں��п����ؽ������ӽ�ϵĹ��ܻ���, �簱�����������[2,3,4], ��ˮ���е��ؽ�������һ����������[5]������ͬ����Ũ��������, ͭ�������Cd2+������Ч�ʴ���Cu2+��Ni2+, ������Ȼ������������ͭ���������������������������ͭ������ǿ[6,7]����ͬ�����Zn2+��Cd2+�������Դ��ڽϴ����, ��״������ (Nostoc punctiforme) ��������1.43 mg/L��Zn2+, ������ (Synechococcus sp.IU625) ������25.00 mg/L��Zn2+[8]��
���������������ؽ������ӵĹ�����Ҳ�������䶾��, ����������Խǿ���ؽ������Ӷ�������������������Խǿ[9]�������о��������, ����ʡ5����� (̫�����������������߱��۲�����������) Cd2+��ƽ������Ũ��Ϊ1.02μg/L, ����̫����Ȫ����Zn2+��Cd2+ȫ��ƽ������Ũ�ȿɴ�31.89μg/L��0.29μg/L, �������Ũ�ȷֱ�Ϊ77.00μg/L��2.20μg/L[10,11]��п�������������������Ԫ��, ������Zn2+��ά������������������л, ��������Zn2+������������, ����ϸ��Ĥ����, ��������½�[12,13]��Cd2+�dz������ؽ�����Ⱦ��֮һ, ���ƻ�����Ҷ����ṹ, ����Ҷ���غϳ�, Ҳ�ܵ��¿�����øʧ������ʱ����Լ�ϸ��Ĥ���˵�[14]�����о��������, 0.05 mg/L��Cd2+����������ͭ����������, ��Cd2+в����, ��ϸ���ᷢ��Ĥ֬��������Ĥ���ľۺ�����, ʹĤϵͳ����, ������Ũ��Cd2+������ϸ�����漰��Χ��״������, ������ϸ�����������ܽ�, ϸ����������, Cd2+��б��դ�� (Scenedesmus obliquus) ������Ӱ�����Ϊ������Ũ�� (<0.10 mg/L) �ٽ�, ������Ũ������[15]��������Ũ�ȵ�Zn2+��Cd2+���ᵼ��ͭ������ϸ����Ҷ����a��Ҷ����b������ܲ��ء��嵨�ص�ɫ�غ����Լ���ϵͳII (PSII) ������ת��Ч�ʽ���, �������嵨����PSII��Ӧ���Ľ�����������[16,17]��
������һЩ���÷������, ˮ���������ؽ�����Ⱦ, �ҳ����д��ģ������ˮ���������˷糡������, ˮ���е������ϸ���ˮ��, ������ˮƽƯ���˶�, ��ʹ�ýϴ��������������ۼ���ij�ض�����[18]����ˮ������ˮƽƯ�ƹ�����, ����Ҳ��Я�������ա�����������һ���˶�, ���ۻ������������ֽ��, �����ڸ��������ʱ��ͷ�, �����»ص�ˮ������, �Ӷ��ı����ʵĿռ�ֲ�[19,20]��ͭ���������γ�����ˮ������Ҫ����, ���ؽ������н�ǿ�ĸ������������, ���о����������������, ��չ��ͬп����Ũ�ȡ�������Ũ����ͭ�����嶾��ѧ����, ���ڲ���ͭ��������ؽ���в�ȵ�������Ӧ���ƺ���������, �Լ���ϸ���ܶȡ���ø���ԡ�PSII������ת��Ч�� (Fv/Fm) �����������绯ø (SOD) ���ԡ���������ø (CAT) ���ԡ�����ȩ (MDA) �����ڲ�ͬп����Ũ�ȡ�������Ũ���µı仯��
����1 �����뷽��
����1.1 �������ּ�����
��������ʹ�õ�ͭ������ (Microcystis aeruginosa) �����й���ѧԺˮ�������о�����ˮ���ֿ⡣��������ǰ, �����������10%��������Һ����3 d, ����ȥ����ˮ��ϴ����ɡ�ͭ��������������ǰ, ����Rippka��[21]�������Ƶ�BG11�������������������120���ѹ��������� (YXQ-LS-50A, ��Ѷ��˾��Ʒ) ���30 min����������̨ (ESCO AC2-4S, ��˼�߹�˾��Ʒ) ��, ��������Һת��������500 ml BG11����������ƿ��, �����ڹ��������� (GZX-300BSH-III, ���繫˾��Ʒ) �н�����������, ���ն�Ϊ (35±5) μmol/ (m2·s) , �ⰵ��Ϊ1��1, �¶ȿ����� (25±1) ��, �����������ж���ҡ����ƿ, �����������ֽ���������
����1.2 ���鴦��
��������������������ͬ, ʹ�ø�����BG11������ (��Zn2+) ��ͭ���������3 d��������[22], �Լ���BG11��������ԭ��Zn2+�ĸ��š���������, ȡ290 ml������BG11����������500 ml��ƿ��, Ȼ��ֱ����5 ml��ͬ����Ũ�ȵ�Zn SO4·7H2O��Cd Cl2·2.5H2O (������) ��Һ, �ټ���5 ml��������������ͭ������Һ, ʹZn2+��Cd2+����Ũ���ݶȾ�Ϊ0 mg/L��0.05 mg/L��0.10 mg/L��0.15 mg/L��0.20mg/L��0.25 mg/L��������ʽϸ���Ǽ��ͭ������ij�ʼ״̬, �Ա�֤�½������״̬��������Ҫ���ݾ��ú����뷢չ��֯ (OECD) ��, ����������ж�������ʱ, ��ij�ʼ�ܶȶ��趨��1 mlԼ1.00×105�� (����������) , �Դ����Ƚϲ�ͬ���ʵĶ��Դ�С[23]���������ʼʱ������ܶ�Ϊ1 ml 1.14×106��, ͭ�����崦�ڶ��������ڡ�Zn2+��Cd2+ÿ������Ũ�Ⱦ���3���ظ�, ��������144 h����ͭ�����������ĵ�0 h��24 h��48 h��72 h��96 h��120 h��144h�ⶨ���ܶȡ���ø���ԡ�PSII������ת��Ч�� (Fv/Fm) , ��ͭ�����������ĵ�0 h��48 h��96 h��144 h�ⶨ���������绯ø���ԡ���������ø���ԡ�����ȩ����, ��ͭ�����������ĵ�144 h�ⶨ��ϸ����Zn2+��Cd2+�ĸ�������
����1.3 ָ��ⶨ
����1.3.1 ���ܶȺ���ø����
������������, ͭ�����������ĵ�0 h��24 h��48 h��72 h��96 h��120 h��144h, �ɼ���Һ2 ml, ʹ��ͨ����Ϊ45μm������ɸ��, ����Һ�ֳ�2�� (�����Լ1 ml) , �ֱ�ⶨ��ø���Ժ���ϸ���ܶȡ���1������Һ�м��������ӫ���� (FDA, Sigma��˾��Ʒ) , ʹFDAŨ��Ϊ25μmol/L, 25�氵������8 min��, ʹ����ʽϸ���� (BD AccuriTMC6, ���˶�-�ҽ�ɭ��˾��Ʒ) �ⶨ��ø����, ����ʽϸ����ֱ�Ӳⶨ��һ������Һ����ϸ���ܶȡ���ʽϸ����Ϊ˫��������:��1����ļ�������Ϊ488 nm, ��������ӫ����ͨ�� (FL1) (530 nm/30 nm) �ռ�FDAӫ��, ������-Ҷ����-�����ʸ���ӫ����ͨ�� (FL3, ��ͨ�˹�Ƭ670 nm) �ռ����ܶ�ӫ��, ��2���⼤������Ϊ640 nm, ��������Ϊ1μl/s, �������Ϊ10μl[24]����ø������ӫ��ǿ�Ȱٷֱȱ�ʾ, ��:���ӫ��ǿ��=�ⶨӫ��ǿ��/��ʼӫ��ǿ��×100%��
����1.3.2 �ܵ����ʺ�����SOD���ԡ�CAT�����Լ�MDA����
������������, ��ͭ�����������ĵ�0 h��48h��96 h��144 h, �ɼ�������Һ��15 ml���Ĺ���, ���䶳̨ʽ���Ļ� (5810R, �����¹�˾��Ʒ) ������ (4�桢10 000 r/min) 10 min, �ռ���ϸ��, ʹ��ϸ����������Ԥ���0.05 mol/L�����λ���Һ (PBS, p H=7.8) ��, ��ԡ�²��ó�����ϸ�������� (X0-1000D, ��ŷ��˾��Ʒ) ������ϸ��, ֱ������������ϸ��, ���� (4�桢10 000 r/min) 10 min, ȡ����Һ, ����ҺΪ�����øҺ����ȡ��ϸ���ڵ�MDAʱ, �������ռ�����ϸ��������10%���������� (TCA) ��, ��������, ����, ȡ����Һ, ����ҺΪ�����MDA��ȡҺ��
���������ʺ������ÿ���˹�������ⶨ[25]��SOD���Բ��õ������� (NBT) �ԭ���ⶨ, ��1 mg��������1 ml��ӦҺ��ʹNBT�ԭ��Ӧ�����ʴ�50%ʱ����Ӧ��SOD��Ϊ1��������λ, SOD���Ե�λΪU/mg��CAT���Բ����������շ�[26]�ⶨ, ��1 mg������1 s�ֽ�1μmol H2O2����Ϊ1��������λ, CAT���Ե�λΪU/mg��MDA������������ͱ����ᷨ (TBA) [27]�ⶨ, ��λΪnmol/mg��
����1.3.3 ���ӫ��
�������ø���ֲ��ӫ���� (PhytoPAM, Walz��˾��Ʒ) �ⶨҶ����ӫ�������ȡ��Һ2 ml, ����Ӧ3 min����в���, ֱ���������϶�ȡPSII������ת��Ч�� (Fv/Fm) ��ֵ, ����ֵ�ɷ�ӳͭ������DZ�ڵ���������ʡ�
����1.3.4 ��ϸ����п���ӵĸ�����
����ͭ�����嶾������������144 h, �ɼ���Һ50 ml�����Ĺ�, �����Ļ� (TDL-40B, ��ͤ��˾��Ʒ) ����4 000 r/min����15min, ��ͭ�������������BG11����������, �㵹��������, ����������ϸ����������ȥ����ˮ��, 10 min��С����ȡ�ϲ���ϸ����Һ, ��ȥ���Ĺܵײ�����, �ٽ���ȡ����Һ��4 000 r/min����15 min, �ռ���ϸ��, �㵹��ϴҺ��������ͭ����������Ĺ������䶳����� (FD-1A-50, ��ҽ����˾��Ʒ) ��, ���������������������ȡ0.05 g�����ķ���ϩ�������, ��5 ml����, ��ҹ, 12 h��������������������, ����, 130������2 h, Ȼ��Ӹ�����2 ml, 160������1 h, ��������180���������, ֱ����Һ����ʱֹͣ����, ��ȴ�����º��5 ml 2%��ϡ����, ֱ��ճ������ܱڵ����������ȫ�ܽ�, ��ȥ����ˮ��������е���Һ�ͳ���ȫ��ת����50 ml����ƿ��, ����, ҡ��, ���ù�ҹ, 12 h������ƿ�е��ϲ���Һ����15 ml���Ĺ���, ���õ����ϵ������������� (ICP-MS, Agilent 7700X, ������˾��Ʒ) �ֱ�ⶨ��ϸ����п���ӵĸ�������
����1.4 ���ݴ�����ͳ��
��������Microsoft Excel 2010��������, ��SPSS21.0�����������е����ط������ (ANOVA) , ������С�����Բ��취 (LSD) ����������ж��رȽ�, ����GraphPad Prism 5������ͼ��
����2 ��������
����2.1 ͭ�������п���ӵĸ�������
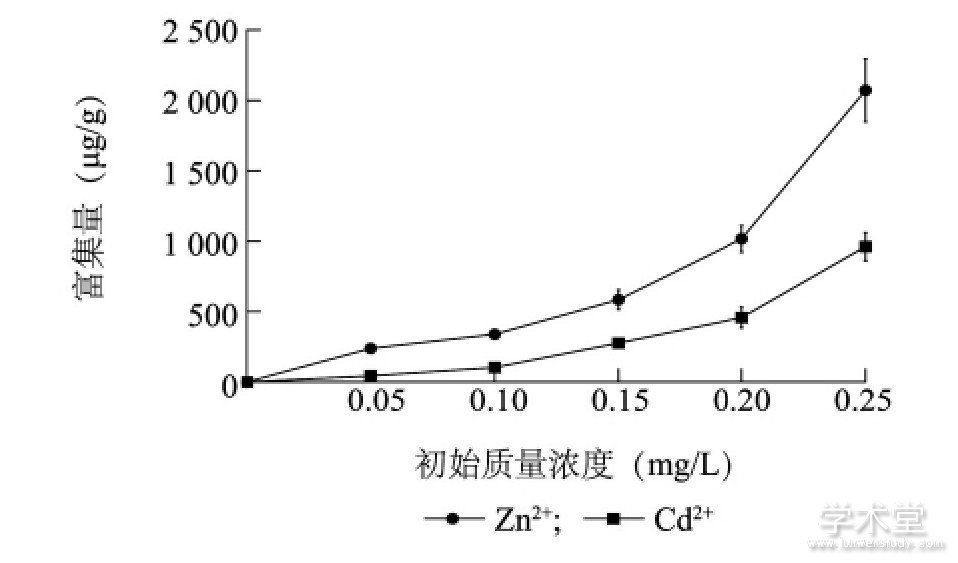
����ͼ1��ʾ, ͭ�������п���ӵĸ���������Zn2+��Cd2+��ʼ����Ũ�ȵ����߶�����Zn2+��Cd2+�ij�ʼ����Ũ����0.05 mg/L������0.25mg/Lʱ, ͭ�������Zn2+��Cd2+�ĸ������ֱ�Ϊ238.27��2 071.45μg/g��43.72��961.67μg/g, 5�������µ�ƽ�������ֱ�Ϊ850.21μg/g��368.57μg/g��ͭ�������Zn2+��Cd2+����һ���ĸ�������, ����ʼ����Ũ����ͬʱ, ��λ���������Zn2+�ĸ��������Ÿ������Cd2+�ĸ����� (P<0.05) ��
����ͼ1 ͭ��������Zn2+��Cd2+�ĸ�����Fig.1 Enrichment of Zn2+and Cd2+in Microcystis aeruginosa
����2.2 Zn2+��Cd2+��ͭ������ϸ���ܶȵ�Ӱ��
����ͼ2��ʾ, �������ڼ�, Zn2+����Ũ��Ϊ0.05mg/L�������ͭ������ϸ���ܶ���ͬʱ�ڶ��������, ����������13.30%��22.90% (P<0.05) , ˵��������Ũ��Zn2+�ܹ��ٽ�ͭ������������Zn2+����48 h, 0.20 mg/L��0.25 mg/L���������ϸ���ܶ����ŵ���ͬ�ڶ����� (P<0.05) , ����144hͭ������ϸ���ܶȷֱ��½���35.01%��70.03%, ����������Ũ��Zn2+��ͭ������ϸ�����������������á�Cd2+����72 h, ������Ũ�ȴ�����ͭ������ϸ���ܶȾ����ŵ���ͬ�ڶ����� (P<0.05) , ����144 h, ����������, 0.05 mg/L��0.10mg/L��0.15 mg/L��0.20 mg/L��0.25 mg/L��Cd2+������ͭ������ϸ���ܶȷֱ��½���10.10%��11.31%��10.63%��22.89%��42.69%����Zn2+��Cd2+��ʼŨ����ͬ�Ҵ��ڵ���0.20 mg/Lʱ, Zn2+��ͭ��������������ñ�Cd2+��������, �����ؽ�����������������Խǿ, ������������������Խǿ�Ľ���һ�¡����о����[20]����, Cd2+��¶48 h�ڶ���ͨС���� (Chlorella vulgaris) �Ķ��Ը���Zn2+, ����ʱ�䱩¶��Cd2+�Ķ���С��Zn2+, ����Zn2+��Cd2+����ͭ�������ЧӦһ�¡�
����ͼ2 Zn2+��Cd2+��ͭ������ϸ��������Ӱ��Fig.2 Effects of Zn2+and Cd2+on the growth of Microcystis aeruginosa
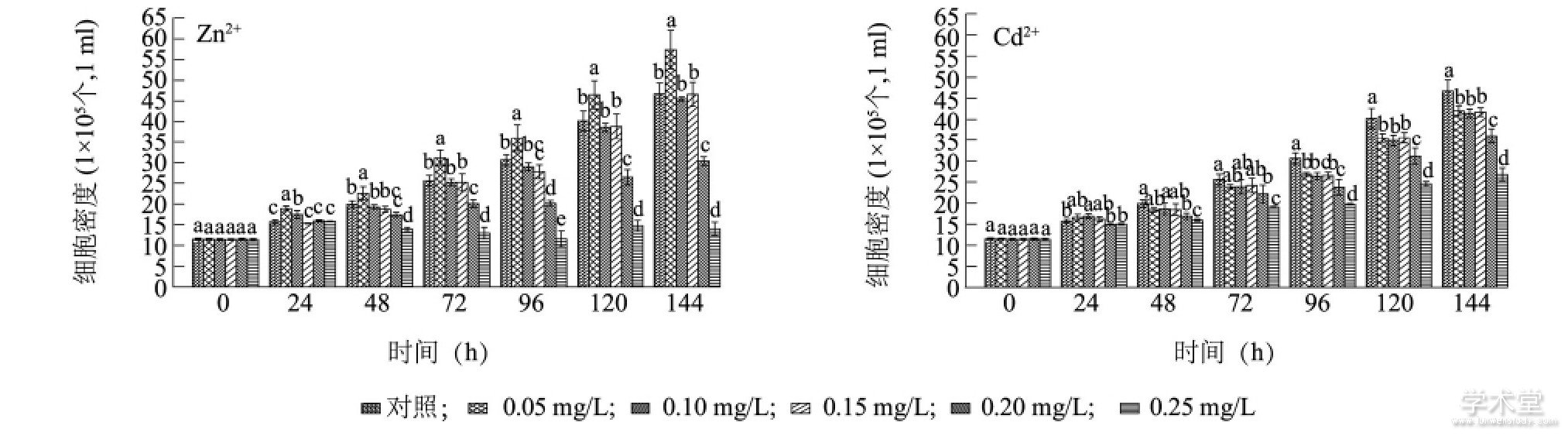
����ͼ�в�ͬСд��ĸ��ʾͬʱ�̲�ͬ����Ũ���ؽ�����������ϸ���ܶȲ������š�
����2.3 Zn2+��Cd2+��ͭ��������ø���Ե�Ӱ��
������ø���Կ��Է�ӳϸ�����³´�л����, �DZȽ����пɿ��Ķ���ѧָ�ꡣ������ӫ���� (FDA) Ⱦ���ܹ���ϸ��Ĥ����øˮ��, ����û��ӫ��, ����ˮ�������Է���ӫ�⡣���, ����ͨ�������Ӧ��ӫ���ź�����ӳ��ϸ���Ļ���[28]��
����ͼ3��ʾ, Zn2+����24 h, 0.05 mg/L����������ӫ��ǿ����������������Բ��� (P>0.05) , ��Zn2+����Ũ�ȸ���0.10 mg/L����������ӫ��ǿ�Ⱦ����Ը��ڶ����顣48 h��, 0.15 mg/L����������ӫ��ǿ�Ȳ����½�, 96 h����ͬ�ڶ������������Բ��� (P>0.05) , ��0.20 mg/L��0.25 mg/L����������ӫ��ǿ��ʼ�����Ÿ���ͬ�ڶ����� (P<0.05) ������48��144 h, 0.20 mg/L Zn2+��0.25mg/L Zn2+����������ӫ��ǿ�ȷֱ���ͬ�ڶ������217.5%��246.5%��280.7%��209.2%��191.3%��481.8%��502.2%��483.5%��320.2%��267.6%��Cd2+����24 h, ������Ũ�ȴ�������ϸ�������ӫ��ǿ�Ⱦ����Ÿ���ͬ�ڶ����� (P<0.05) ������48 h��, 0.05 mg/L��0.10 mg/L����������ӫ��ǿ����ͬ�ڶ������������Բ���, ��0.15 mg/L��0.20mg/L��0.25 mg/L����������ӫ��ǿ��ʼ�����Ÿ���ͬ�ڶ����� (P<0.05) , ����96 hʱ�ﵽ���, �ֱ��Ƕ��������ӫ��ǿ�ȵ�272.8%��537.2%��757.3%���ؽ�������48 h, ����Ũ��Ϊ0.25 mg/L��Zn2+��Cd2+����������ӫ��ǿ�ȷֱ�������24 hʱ���ӫ��ǿ�ȵ�206.1%��118.7%��
����ͼ3 Zn2+��Cd2+��ͭ��������ø���Ե�Ӱ��Fig.3 Effects of Zn2+and Cd2+on the esterase activity of Microcystis aeruginosa
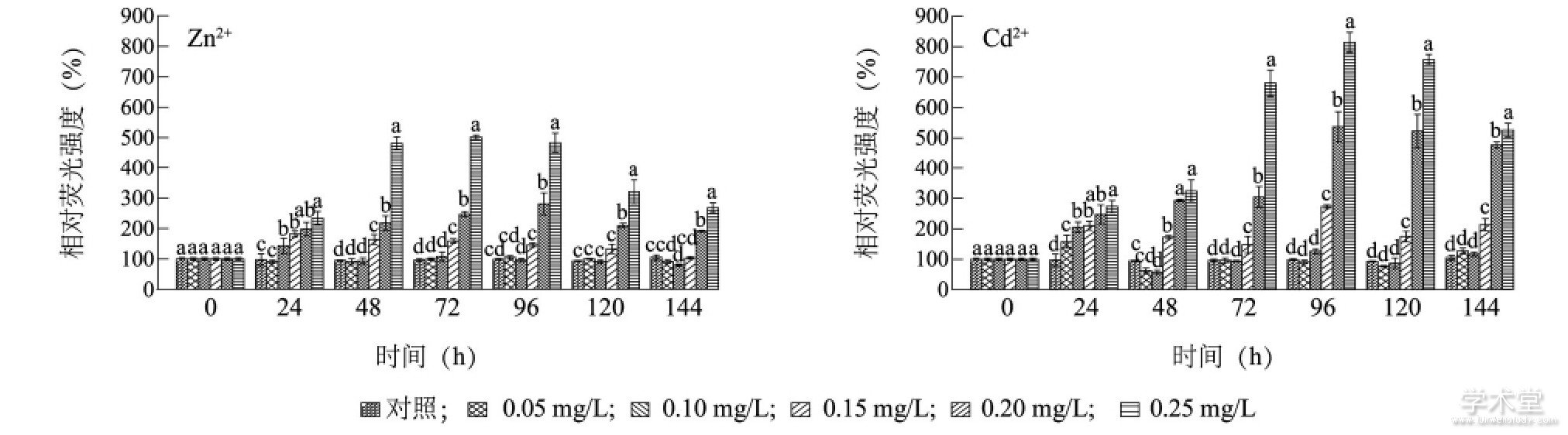
����ͼ�в�ͬСд��ĸ��ʾͬʱ�̲�ͬ����Ũ���ؽ�����������ϸ�����ӫ��ǿ�Ȳ������š�
����2.4 Zn2+��Cd2+��ͭ�����������õ�Ӱ��
����PSII������ת��Ч���ֳ�PSII�Ĺ⻯ѧ������Ӳ���, �ǰ���Ӧ��PSII�����⻯ѧЧ��, ��ӳ�˹����������DZ�ڵ��������������, ����Ϊֲ���������Ƿ������������ָ��[29]��
����ͼ4��ʾ, ������120 h��, Zn2+����Ũ�ȵ���0.25 mg/L��������ϸ��Fv/Fmֵ������������, 120 h�����ȶ�, ���ҳ���0.20 mg/L�������Fv/Fmֵ���ŵ����������� (P<0.05) , �������첻���š�Zn2+Ũ��Ϊ0.25 mg/L�������Fv/Fmֵ������24 h�����ŵ���ͬ�ڶ����� (P<0.05) , �����ڼ������ʱ�̵�Fv/Fmֵ֮�������Ų���, 72h��, 0.25 mg/L�������Fv/Fmֵ�����ŵ���ͬ�ڸ�����Ũ�����Fv/Fmֵ (P<0.05) ��Cd2+��ͭ������Fv/Fmֵ��Ӱ����Zn2+����, �����ڼ�, ������Ũ�����Fv/Fmֵ��120 hǰ��������, 120 h���ȶ���48 h��, Cd2+����Ũ��Ϊ0.25 mg/L�������Fv/Fmֵ���ŵ���ͬ�ڸ�����Ũ���� (P<0.05) , ������ͬ����Ũ��Zn2+в������ϸ����Fv/Fmֵ��
����ͼ4 Zn2+��Cd2+��ͭ�����������õ�Ӱ��Fig.4 Effects of Zn2+and Cd2+on the photosynthesis of Microcystis aeruginosa
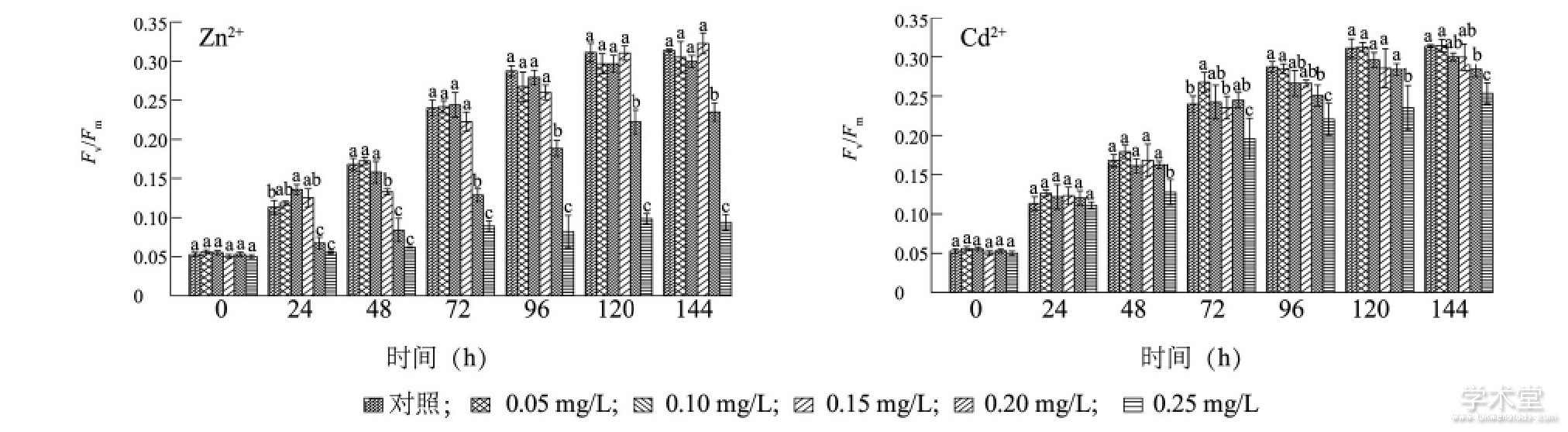
����ͼ�в�ͬСд��ĸ��ʾͬʱ�̲�ͬ����Ũ���ؽ�����������ϸ��Fv/Fmֵ�������š�
����2.5 Zn2+��Cd2+��ͭ�����忹�����Ե�Ӱ��
����ͼ5��ʾ, Zn2+��¶48 h, ������Ũ�ȴ�������ϸ����SOD������ͬ�ڶ���������������Բ��� (P>0.05) ��0.10 mg/L��0.25 mg/L Zn2+�������CAT���Էֱ����ŵ��ں���ͬ�ڶ����� (P<0.05) , ����������Ũ�ȵ�Zn2+�̼�����ϸ����CAT���Ե����ߡ�����96 h, 0.15 mg/L��0.20 mg/L Zn2+�������SOD�������Ÿ���ͬ�ڶ����� (P<0.05) , ��0.25 mg/L Zn2+�������SOD�������ŵ���ͬ�ڶ����� (P<0.05) ������96 h, 0.05 mg/L��0.10 mg/L Zn2+�������CAT�������ŵ���ͬ�ڶ����� (P<0.05) , 0.15��0.25 mg/L�������CAT��������������, �������Բ��� (P>0.05) , ����������Ũ��Zn2+��һ���̶�����������ϸ����CAT���ԡ�����144 h, 0.15��0.25 mg/L Zn2+��������ϸ����SOD�������Ÿ���ͬ�ڶ����� (P<0.05) , 0.20 mg/L��0.25 mg/L�������CAT����Ҳ���Ÿ���ͬ�ڶ����� (P<0.05) , ����������Ũ�ȵ�Zn2+��ʱ�䱩¶��̼�����ϸ��������ø���Ե����ߡ�
��������48 h, 0.10 mg/L Cd2+��������ϸ����SOD�������Ÿ���ͬ�ڶ����� (P<0.05) , ��0.05��0.25 mg/L�������CAT����Ҳ���Ÿ���ͬ�ڶ����� (P<0.05) ��Cd2+��¶96 h, 0.05 mg/L�������SOD�������ŵ���ͬ�ڶ����� (P<0.05) , 0.05��0.15 mg/L�������CAT����Ҳ���ŵ���ͬ�ڶ����� (P<0.05) , ��0.25 mg/L�������CAT�������Ÿ���ͬ�ڶ����� (P<0.05) ������144 h, ��Cd2+��������ϸ����SOD���Ծ����Ÿ���ͬ�ڶ����� (P<0.05) , ��0.25 mg/L Cd2+�������SOD�������, 0.25 mg/L Cd2+�������CAT�������Ÿ���ͬ�ڶ����� (P<0.05) , ����������Ũ��Cd2+���������ϸ����SOD��CAT���ԡ���Zn2+��в������, ��ϸ����MDA�IJ������ֳ�һ���ļ���ЧӦ, ���ؽ�������Ũ��Խ��, MDA�ĺ���Խ��, ��ͬʱ�̸�����Ũ�� (0.20��0.25 mg/L) Cd2+������ϸ��������MDA��������ͬ����Ũ��Zn2+������ϸ��������MDA������
����ͼ5 Zn2+��Cd2+��ͭ������SOD���ԡ�CAT���Ժ�MDA������Ӱ��Fig.5 Effects of Zn2+and Cd2+on SOD activity, CAT activity and MDA content of Microcystis aeruginosa
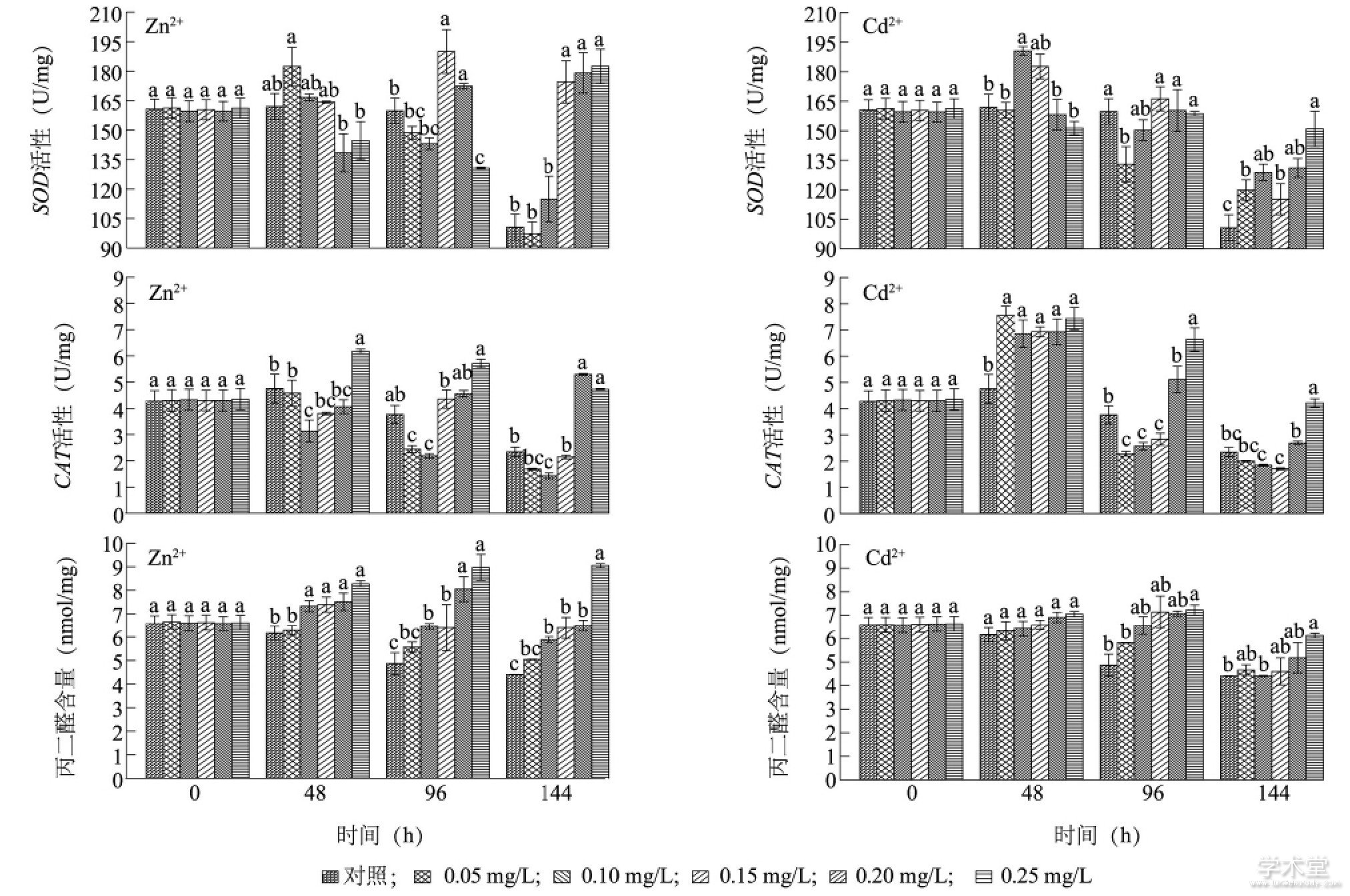
����ͼ�в�ͬСд��ĸ��ʾͬʱ�̲�ͬ����Ũ���ؽ�����������ϸ��SOD���ԡ�CAT���Ժ�MDA�����IJ������š�
����3 ����
��������ϸ�����ϴ��ڴ����İ������ǻ�����������������ȹ�����, �����ؽ������ӽ��, ���ؽ�������ǿ�Ĺ�����������ˮ�����Ӱ뾶�Լ����Ӳ�ṹ���ԵȾ��ǵ�������Բ�ͬ�ؽ����������Լ������������ڲ������Ҫ����[30,31,32]��п���Ӿ�Ϊ���ɽ���Ԫ��, ������ͬ���������λ����, ���ڱ��о���ͭ�������п���ӵĸ�����֮������߳�ʼ����Ũ�ȵ����Ӷ��½�, ��ֵ��4.5���½���2.2��Fisher[9]��Ϊ, Zn2+��Cd2+���뺬�����������Ž��, Zn2+ˮ�ϰ뾶��Cd2+С, ʹ��ͭ�������Zn2+������ǿ��Cd2+, ������ǵ���ͭ�������Zn2+�и��߸����Ե���Ҫԭ����贵�[15]���о��������, ����Cd2+��ʼ����Ũ�ȵ�����, ͭ�������б��ɺ���Cd2+�ĸ�����������, ��Cd2+����Ũ��Ϊ0.20 mg/Lʱ, ͭ�������б��ɺ��ϸ���ڵ�Cd2+�����ֱ�Ϊ546.00μg/g��769.00μg/g, �뱾�о���ͬ��ʼCd2+����Ũ�ȴ����µ�ͭ������Cd2+������ (457.52μg/g) �����൱����Zn2+��Cd2+����Ũ�ȳ�������0.25 mg/Lʱ, ������ϸ��Ĥ���ܵ�����, ��������, ϸ���ȱ��������, ���汩¶����������ؽ�����ϵĻ��Թ�����, �����ܸ���һ�������ؽ���, �����帻���ؽ����Ļ�����ϸ������ת��Ϊ����[33]��
����̼����ø��ˮ���е���̼ת��Ϊ̼ˮ������, ��п��̼����ø����Ҫ���, ������Zn2+����߹������ǿ��, �ٽ���������, ����Ũ�ȵ�Zn2+�谭Ҷ���廹ԭ��øII�γ�, ���º���ֽ�[34]�������� (Synechococcus sp.IU 625) �����ܸߴ�25 mg/L��Zn2+, ��50 mg/L��Zn2+ʹ������ϸ��ɥʧ����, ��ϸ�����Ѳ���ȫ, ϸ�������������, ��ϸ������ֱ�ӵĶ�������[8]��Cd2+��ͭ�����������ķDZ���Ԫ��, ��Cd2+���۵�һ����ʱ���ͭ�����������������, ������贵�[15]���о����һ�¡���Cd2+����Ũ�ȴﵽ25 mg/Lʱ, ��������ϸ�������������ʽӽ�50%, �����嵨�����ʵ�����Ũ�������½�[35]������Cd2+����Ũ�ȵ�����, ��βդ�� (Scenedesmus quadricanda) ϸ����Ҷ����a��Ҷ����b������ܲ��ص�ɫ���Լ������Էӵĺ����½�, �������������Ӻ�������ĺ�������[17,36]���ɼ�, ���ڲ�ͬ�������̬��ϸ��Ĥ��ϸ���ڵ������ṹ���ڲ���, �������ͬ���ؽ�������ֳ���ͬ�������ԡ�
������������24 h��, Zn2+��Cd2+����Ũ�ȷֱ�ﵽ0.20 mg/L��0.15 mg/Lʱ, ͭ���������ø���Ծ����Ÿ���ͬ�ڶ�����, ����ϸ���ܶ�ʼ�յ���ͬ�ڶ�����, ����Zn2+��Cd2+��в������������ͭ�����������, ��ȴ��ʹ��ϸ����ø�������ߡ���ø���Ե����߱�����ϸ���³´�л���ǿ, һ����������ؽ���������ϸ��Ĥ����֬��л, �Ӷ��̼��˰��ڵ���ø, �����ṩ�����������ά��ϸ������, ���ؽ���ѹ���¶�ʱ��������������ұ���������һ��Ӧ����ʽ[37,38]����һ����, �����Ǹ�����Ũ���ؽ�����ʹ��ϸ��Ĥ������״̬�����ı�, Ĥϵͳ��л��������, ������ͭ�������������ֳ[38], �Ӷ�ʹ��ϸ���ܶȽ��͡����о�ָ��, ��Zn2+��Cd2+Ũ�ȷֱ�ﵽ0.15μmol/L��0.12μmol/Lʱ, ��ͷ״α������ (Pseudokirchneriella subcapitata) ����ø���Ծ��ܵ���������, ��Zn2+��Cd2+Ũ�ȷֱ�ﵽ0.60μmol/L��0.50μmol/Lʱ, ��ø���������ʷֱ�ﵽ17%��28%, ������ϸ����п����в�ȵ������Խϵ�[39]����Cu2+����ͭ������1 h��, ��ϸ����ø������������, Cu2+����Ũ��Ϊ13��101μg/Lʱ, ������ø���������ߺ�, ����ϸ���ڵ���ø�ܵ��˴̼�[40]��Lage��[41]��Ϊ, ���ִ̼���������ΪCu2+�Ľ���ʹϸ�����������µ���, ��������ϸ����p H�������µ�, Ҳ�п�����Cu2+в����, �����ʱ���ڽ������ұ�����һ��Ӧ����ʽ��
����Zn2+��Cd2+в����ͭ�������Fv/Fmֵ����ϸ���ܶȵı仯��������, ��������Ũ��Zn2+��Cd2+в�Ⱦ��ܽ�����ϸ���ܶȺ�Fv/Fmֵ, Fv/Fmֵ�Ľ���˵��PSII������������, ���Ӵ��ݹ�������, ͭ������Ĺ�������ܵ�������[42], ����ʹͭ������������ܵ�����, ��ϸ���ܶȽ��͡�����24 h��, ��Zn2+��Cd2+����Ũ�ȷֱ�ﵽ0.20mg/L��0.25 mg/Lʱ, ͭ�������Fv/Fmֵ�����ŵ���ͬ�ڶ�����, ˵����Ĺ�������ܵ�������, �ؽ�������Ũ��Խ��, ͭ�����������������Ƶij̶�Խ��, ������ͬ������Ũ�� (0.25 mg/L) ��в����, Zn2+��ͭ�������������������ñ�Cd2+���ԡ�пŨ�ȸ���23μmol/Lʱ, �����ƴ��������Ĺ�����ú�����[43]��1 mmol/L��Zn2+��Cd2+���ڶ��� (0.5��2.0 h) ��Ѹ�ٽ��͵���С����S-39 (Chlorella pyrenoidosa Chick S-39) �Ĺ�Ϸ������ʺ�Fv/Fmֵ, ʹPSII��ϵ��Ӵ��ݹ����ܵ�����[[44]]��Zn2+��Cd2+в�Ⱦ��ܽ�����ͨС���� (Chlorella vulgaris) ��������, ����¶96 h��, PSII�⻯ѧ������Ӳ���������Լ10%, ������ǵ�Ũ���ؽ����̼�ЧӦ�Ľ��[[22]]��
����������������������л������, ���������ɻ��IJ�����������ڶ�̬ƽ��״̬, ���������ܵ���������в��ʱ, ϸ���ڻ���������������ɻ�, SOD��CAT�ȿ�����ø��������ϸ���ڶ��������ɻ��ķ�����ϵ, �����ɻ��ɱ�������ø���շֽ�ΪH2O��O2, �Ӷ���ֹ�����ϸ�����˺�����������ø����ϵͳ�ܵ��ƻ�ʱ, �������Ļ��ۻ�����Ĥ��ֲ�����֬����Ĺ�����, ����MDA��Ĥ֬�ʹ�������������[45]��������Ũ��Zn2+��Cd2+�ɶ�ͭ�����������������������, ����ϸ����ͨ��SOD��CAT���������������ɻ�, �Ӷ��ﵽ������ϸ����Ŀ�ġ���Zn2+��Cd2+����Ũ�ȴﵽ0.25 mg/Lʱ, ��������ʱ����ӳ�, CAT���������ߺ�, ����ϸ��ǰ���ܵ������յ�����, ��������������������ɻ�, ����ʹø�ṹ��ƻ�, ����Ĥ֬��������Ĥ���ۺ�����, ʹĤϵͳ����, MDA��������, ����ʹ��ϸ�������ܵ�ǿ��������������, ʹ��ϸ���ܶȽ��͡�Choudhary��[46]�о��˲�ͬ����Ũ��Zn2+ (0.05��0.20 mg/L) ��������-S5 (Spirulina platensis-S5) �Ķ�������, ����������-S5ϸ���ڵ�MDA������SOD�����Լ������Ậ������������, �Ҹ�����������ٷֱ���0.05 mg/L Zn2+������38.4%���ӵ�0.20 mg/L Zn2+������93.3%, MDA���������ӱ���������-S5��Zn2+в�����γ��˻��������ɻ�, ��SOD���Ժ����Ậ�����������, ��ϸ���ܵ������յ����ú�ϸ������������ɻ��Ļ��ƿ�ʼ�������á�
�����ο����ף�
����[1]������, ������, ����Ӣ, ��.����������������������ͭ��п����������������̬���շ���[J].�Ϸ�ũҵѧ��, 2018, 49 (9) :1865-1872.
����[2] PEREIRA S, MICHELETTI E, ZILLE A, et al. Using extracellu-lar polymeric substances (EPS) -producing cyanobacteria for thebioremediation of heavy metals:do cations compete for the EPSfunctional groups and also accumulate inside the cell?[J]. Micro-biology, 2011, 157 (2) :451-458.
����[3]��ƻ, ������.����ϸ�����ؽ���������Լ��俹�Ի���[J].������ѧ�뼼��, 2009, 32 (1) :84-89.
����[4]�¾���, ����Ө, �����, ��.������������̶�����Ⱦ�������������Ч��[J].����ũҵ��ѧ, 2018, 46 (20) :320-324.
����[5] CHAKRABORTY N, BANERJEE A, PAL R. Biomonitoring of lead, cadmium and chromium in environmental water from Kolkata, Northand South 24-Parganas using algae as bioreagent[J]. Journal of AlgalBiomass Utilization, 2011, 2 (3) :27-41.
����[6]���ľ�, ���, ����, ��.����ˮ������Դ������:�����ؽ�������Cu2+��Cd2+��Ni2+��ʵ���о�[J].������ѧ, 2014, 26 (3) :417-422.
����[7] SINGH S, PRADHAN S, RAI L C. Comparative assessment ofFe3+and Cu2+biosorption by field and laboratory-grown Microcystis[J]. Process Biochemistry, 1998, 33 (5) :495-504.
����[8] NEWBY J R R, LEE L H, PEREZ J L, et al. Characterization ofzinc stress response in cyanobacterium Synechococcus sp. IU 625[J]. Aquatic Toxicology, 2017, 186:159-170.
����[9] FISHER N S. On the reactivity of metals for marine phytoplankton[J]. Limnology and Oceanography, 1986, 31 (2) :443-449.
����[10]��ΰ, �����, �ƴ���, ��.����ʡ������ˮ���ؽ����ļ����ȽϷ���[J].������ѧ, 2016, 28 (3) :494-501.
����[11] SU Y, LIU H, YANG J. Metals and metalloids in the water-bloom-forming cyanobacteria and ambient water from nanquan coast ofTaihu lake, China[J]. Bulletin of Environmental Contaminationand Toxicology, 2012, 89 (2) :439-443.
����[12] SUMAN T Y, RAJASREE S R R, KIRUBAGARAN R. Evaluationof zinc oxide nanoparticles toxicity on marine algae Chlorella vul-garis through flow cytometric, cytotoxicity and oxidative stress a-nalysis[J]. Ecotoxicology and Environmental Safety, 2015, 113:23-30.
����[13]���, ��־��.пв�ȶ�С���忹����ø���������Ӱ��[J].��̬ѧ��, 2012, 32 (22) :7117-7123.
����[14] FERNANDEZ-GARC?A N, MART?M C, JIMENEZ A, et al.Sub-cellular distribution of glutathione in an Arabidopsis mutant (vtc1) deficient in ascorbate[J]. Journal of Plant Physiology, 2009, 166 (18) :2004-2012.
����[15]���, �ƺ�, ����ͮ, ��.�Ӷ�ͭ�������б��դ��Ķ���ЧӦ[J].�й�������ѧ, 2014, 34 (2) :478-484.
����[16] XU K, LI Z K, QIU B S, et al. Different responses to high lightstress of toxic and non-toxic Microcystis aeruginosa acclimated un-der two light intensities and zinc concentrations[J]. Toxicological&Environmental Chemistry, 2013, 95 (7) :1145-1156.
����[17]������, �̽��, Ф��¶, ��.��βդ����ؽ�����в�ȵ���Ӧ[J].ũҵ������ѧѧ��, 2015, 34 (4) :633-638.
����[18]����, ���ٻ�, �߿���, ��.̫������ˮ����Ԥ����Ԥ���Ԥ����������ʵ��[J].������ѧ, 2009, 21 (3) :314-328.
����[19]������, ������, Ҷ��, ��.��ϸ�����Լ�Ӫ����ˮƽ��ͭ�����������ӵ�Ӱ���о�[J].�����Ƽ�, 2013, 26 (3) :1-4.
����[20]����, �����, ����, ��.��������ۼ����ؽ���Ǩ���ͷŵ�Ӱ��[J].������ѧ, 2017, 29 (3) :558-566.
����[21] RIPPKA R, DERUELLES J, WATERBURY J B, et al. Genericassignments, strain histories and properties of pure cultures of cya-nobacteria[J]. Microbiology, 1979, 111 (1) :1-61.
����[22] OUYANG H L, KONG X Z, HE W, et al. Effects of five heavymetals at sub-lethal concentrations on the growth and photosynthe-sis of Chlorella vulgaris[J]. Chinese Science Bulletin, 2012, 57 (25) :3363-3370.
����[23]ŷ������, ������, �Ϋ^˪, ��. Cu2+����ͨС����Ĺ�϶���:��ʼ���ܶȵ�Ӱ��[J].��̬����ѧ��, 2011, 6 (5) :499-506.
����[24]���, �����, ��Ƽ��, ��.����Һ�����ļ��Զ���ЧӦ[J].��ȫ�뻷��ѧ��, 2016, 16 (1) :381-386.
����[25] BRADFORD M M. A rapid and sensitive method for the quantita-tion of microgram quantities of protein utilizing the principle of pro-tein-dye binding[J]. Analytical Biochemistry, 1976, 72 (1/2) :248-254.
����[26] BEAUCHAMP C, FRIDOVICH I. Superoxide dismutase:im-proved assays and an assay applicable to acrylamide gels[J]. Ana-lytical Biochemistry, 1971, 44 (1) :276-287.
����[27] NAKANO Y, ASADA K. Hydrogen peroxide is scavenged by a-scorbate-specific peroxidase in spinach chloroplasts[J]. Plant andCell Physiology, 1981, 22 (5) :867-880.
����[28]��ִΰ, ����÷, ���ľ�, ��.�����ζ���ͨС����������Լ��������Ե�Ӱ��[J].������ѧ, 2016, 37 (6) :2158-2163.
����[29] KUMAR K S, DAHMS H U, LEE J S, et al. Algal photosyntheticresponses to toxic metals and herbicides assessed by chlorophyll afluorescence[J]. Ecotoxicology and Environmental Safety, 2014, 104 (2) :51-71.
����[30] VERHEYEN L, VERSIEREN L, SMOLDERS E. Natural dis-solved organic matter mobilizes Cd but does not affect the Cd up-take by the green algae Pseudokirchneriella subcapitata (Korschik-ov) in resin buffered solutions[J]. Aquatic Toxicology, 2014, 154:80-86.
����[31] SHENG P X, TING Y P, CHEN J P, et al. Sorption of lead, cop-per, cadmium, zinc, and nickel by marine algal biomass:charac-terization of biosorptive capacity and investigation of mechanisms[J]. Journal of Colloid and Interface Science, 2004, 275 (1) :131-141.
����[32] GONZ?LEZ-D?VILA M. The role of phytoplankton cells on thecontrol of heavy metal concentration in seawater[J]. Marine Chem-istry, 1995, 48 (3/4) :215-236.
����[33] DA COSTA A A, DE FRANCA F P. Cadmium uptake by Spirulinamaxima:toxicity and mechanism[J]. World Journal of Microbiolo-gy and Biotechnology, 1998, 14 (4) :579-581.
����[34]������, �Ź�ɭ, ����ɽ, ��.п��2�ֵ�ˮ����������ֳ��Ӱ��[J].����ֲ��ѧ��, 2006, 26 (8) :1722-1726.
����[35]����, �����, ���ٱ�, ��. Cd2+в�ȶ��������������������Լ��嵨��������Ũ�ȵ�Ӱ��[J].��̬����ѧ��, 2007, 16 (3) :767-770.
����[36] KOV?��CIK J, KLEJDUS B, ?TORK F, et al. Comparison of meth-yl jasmonate and cadmium effect on selected physiological parame-ters in scenedesmus quadricauda (chlorophyta, chlorophyceae) [J]. Journal of Phycology, 2011, 47 (5) :1044-1049.
����[37]����, ֣��÷, ������, ��.ȫ�������б��դ��ϸ��Ĥ����Ӱ���о�[J].��̬����ѧ��, 2010, 5 (4) :537-542.
����[38] JIAO Y, OUYANG H L, JIANG Y J, et al. Effects of phosphorusstress on the photosynthetic and physiological characteristics ofChlorella vulgaris based on chlorophyll fluorescence and flow cyto-metric analysis[J]. Ecological Indicators, 2017, 78:131-141.
����[39] MACHADO M D, SOARES E V. Use of a fluorescence-based ap-proach to assess short-term responses of the alga Pseudokirchneriel-la subcapitata to metal stress[J]. Journal of Applied Phycology, 2015, 27 (2) :805-813.
����[40]����, ����, ������, ��.Ӧ����ʽϸ�������о�ͭ����ϸ��Ĥ�����Լ�֬ø���Ե�Ӱ��[J].Ӧ���뻷������ѧ��, 2006, 12 (5) :706-709.
����[41] LAGE O M, SANSONETTY F, O'CONNOR J E, et al. Flow cyto-metric analysis of chronic and acute toxicity of copper (II) on themarine dinoflagellate Amphidinium carterae[J]. Cytometry, 2001, 44 (3) :226-235.
����[42] MURCHIE E H, LAWSON T. Chlorophyll fluorescence analysis:aguide to good practice and understanding some new applications[J]. Journal of Experimental Botany, 2013, 64 (13) :3983-3998.
����[43] XU K, JUNEAU P. Different physiological and photosynthetic re-sponses of three cyanobacterial strains to light and zinc[J]. Aquat-ic Toxicology, 2016, 170:251-258.
����[44] PLEKHANOV S E, CHEMERIS Y K. Early toxic effects of zinc, cobalt, and cadmium on photosynthetic activity of the green algaChlorella pyrenoidosa Chick S-39[J]. Biology Bulletin of the Rus-sian Academy of Sciences, 2003, 30 (5) :506-511.
����[45] COLL?N J, PINTO E, PEDERSEN M, et al. Induction of oxida-tive stress in the red macroalga Gracilaria tenuistipitata by pollutantmetals[J]. Archives of Environmental Contamination and Toxicolo-gy, 2003, 45 (3) :337-342.
����[46] CHOUDHARY M, JETLEY U K, KHAN M A, et al. Effect ofheavy metal stress on proline, malondialdehyde, and superoxidedismutase activity in the cyanobacterium Spirulina platensis-S5[J].Ecotoxicology and Environmental Safety, 2007, 66 (2) :204-209.





