
全世界干旱、半干旱荒漠地区占到陆地面积的33%~40%,在这些地区有限的水分承载力限制了大多维管植物的生存,然而生物结皮却能够广泛分布,甚至在一些地区占到生物覆盖的70%以上.生物结皮是由藻类、地衣、苔藓、真菌及异养细菌等生物组分胶结土壤颗粒在地表形成的一层易剥离的复合生物土壤层,它们在荒漠地区的土表稳定、营养改善、水分循环等方面起着非常重要的作用.同时,促进生物结皮的生长发育也被认为是抑制沙尘、改善荒漠生境最有效的方法之一.正是由于其重要的生态功能以及对荒漠环境独特的贡献,生物结皮在近些年来引起了国内外众多研究人员的广泛关注.
生物结皮作为荒漠地区典型的生命存在形式,自身一直处在不断的发育演替过程当中.在结皮的发育演替过程中,根据优势种类的不同,一般将生物结皮划分为藻结皮、地衣结皮和藓结皮这3种主要的演替类型.藻结皮的优势类群为丝状蓝藻,但随着结皮的发育,地衣、藓类逐渐成为结皮表面的优势类群,结皮发育演替至地衣结皮或藓结皮.随着结皮的发育演替,结皮中生物群落结构不断变化,而不同的生物类群势必会影响结皮的生态功能.已有研究证实,不同发育演替阶段生物结皮在固碳、固氮效率、抗压强度、土壤质地、土壤孔隙度等方面存在明显差别.然而目前对于导致结皮这些生态功能及理化特性方面差异的内在机制并没有进行深入的研究.尤其在结皮的发育演替过程中,微生物组成及生物量变化方面的研究非常缺乏.而开展此方面的研究,将有助于从本质上揭示不同发育演替阶段生物结皮在生态功能、理化特性及各属性之间的差异.
1、材料与方法
1.1试验区概况与样品采集
试验区位于腾格里沙漠东南边缘的沙坡头地区(37°32'N,105°02'E,海拔1339m),处于阿拉善高原荒漠与荒漠草原过渡地带,属典型的大陆性季风沙漠气候.该地区年平均气温9.6℃,平均降雨量为180mm,降雨主要集中在7~9月;空气平均相对湿度为40%,年平均蒸散量大于3000mm,主要集中在生长季节(5~9月).该地区的年平均风速为2.6m·s-1,大于5m·s-1的起沙风每年有200d左右.包兰铁路从这里经过,为确保铁路正常运行,从1956年开始该地区进行人工植被恢复.经过长期的演替,该地区目前生态环境得到了改善,大量的生物结皮在沙土表面形成.
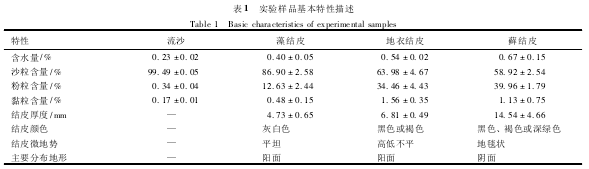
实验所用结皮及流沙样品于2010年6月采集于沙坡头地区的人工植被恢复区,样品基本特征见表1.结皮及流沙样品用环刀和无菌铲采集后装于无菌培养皿中,Parafilm膜封口,所有样品快速运回实验室后一个月内进行相关的实验分析.
1.2叶绿素a含量的测定
结皮中的光合生物量用叶绿素a表示.叶绿素a测定时,将同一样品分为两份,一份直接测定结皮中总的叶绿素a含量,另一份则先去除地衣叶状体和藓类“茎叶”部分,之后再测定其中的叶绿素a含量.测定时,样品先用研钵充分研磨,之后取1g左右土样加入10mL丙酮在4℃提取过夜,叶绿素a含量按照Carcia-Pichel等的三色公式进行计算.
1.3优势藻类的直接观察鉴定
结皮中优势藻类的确定利用结皮-水混合液在显微镜下直接观察鉴定并计数.首先将结皮样品过150目筛,然后取0.1g过筛结皮样品加入10mL蒸馏水,混匀后在暗处放置12h.之后取0.1mL结皮-水混合液在显微镜下观察,根据藻类的显微形态特征对其鉴定,同时计算各种藻的生物体积,计算方法参照文献的方法.
1.4异养微生物数量的确定
将结皮样本在无菌条件下过150目筛,然后将过筛后的结皮样品用无菌水稀释成不同浓度梯度的结皮-水混合液,混匀后在暗处放置12h.之后在每一稀释浓度的结皮-水混合液中分别取3份0.1mL结皮-水混合液涂布于牛肉膏蛋白胨培养基、马丁氏培养基和高氏I号培养基培养,用于培养结皮中的细菌、真菌和放线菌.培养细菌的平板置于37℃培养,培养放线菌和真菌的平板置于28℃培养.培养3~5d后进行菌落计数.
1.5结皮理化特性测定
结皮的理化特性包括含水量、质地组成(其中沙粒、粉粒、黏粒的直径分别为>0.05mm、0.005~0.05mm、<0.005mm)、有机碳、总氮及总磷含量按照文献中所描述的方法进行测定.结皮厚度用游标卡尺进行测定;结皮pH和电导率在结皮5∶1水土比浸出液中用YSI-Proplus型多参数水质分析仪进行测定;结皮K+、Na+、Ca2+、Mg2+、Mn2+含量在M3土壤浸提液中用原子吸收分光光度法进行测定.
1.6统计分析
本研究中不同演替阶段生物结皮中叶绿素a含量、藻类生物体积及异养微生物菌落数量之间的方差分析利用One-WayANOVA进行分析;藻类生物体积及异养微生物菌落数量与结皮理化特性之间的相关性分析用BivariateCorrelations进行分析.所有的数据分析在SPSS13.0软件上进行.
2、结果与分析
2.1生物结皮的发育演替
藻结皮以蓝藻为优势群落,尤其是丝状蓝藻,如具鞘微鞘藻(Microcoleusvaginatus)、爪哇伪枝藻(Scytonemajavanicum).藻结皮较薄,平坦、易碎,表面颜色较浅,藻类盖度一般>80%.地衣结皮中以胶质状地衣为主,如坚韧胶衣(Collematenax).地衣结皮厚度增加,表面较粗糙,表面颜色加深,地衣叶状体分布在结皮表面,且此时藓类很少出现,干燥时地衣最大盖度可达到80%,湿润时则覆盖整个结皮表面.藓结皮中以真藓和对齿藓为主,如银叶真藓(Bryumargenteum)、黑对齿藓(Didymodonnigrescens)等.藓结皮厚度可达到15mm,表面被地毯状的藓类植物体所覆盖,盖度有时可达到100%.调查显示该试验区生物结皮一般按照“藻结皮?→地衣结皮?→藓结皮”的模式进行发育演替.在一些情况下,藓类也可直接在藻结皮上萌发生长,从而从藻结皮直接演替到藓结皮(图1).

2.2结皮中光合生物生物量变化
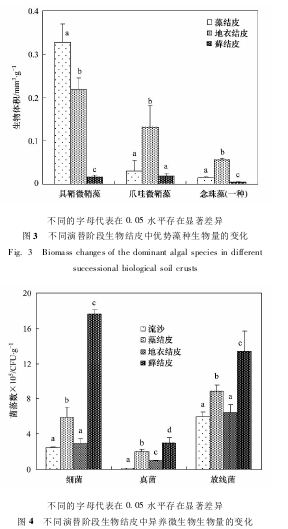
作为结皮中重要的生物类群,光合生物是结皮有机碳的主要贡献者,在结皮的生长、发育、演替、恢复过程中都起着重要作用.本研究同样发现,随着结皮的发育演替,结皮中的光合生物量呈逐步增加的趋势(图2).当把地衣的叶状体和藓类的“茎叶”部分去除后,结皮中的光合生物则仅剩下自由生长的藻类.由图2结果显示,自由生长的藻类在藻结皮、地衣结皮到藓结皮的演替过程中呈下降的趋势.然而尽管从藻结皮到地衣结皮,自由生长的藻类生物量有所下降,但总的藻类生物量却呈逐渐增加.这说明,当演替到地衣结皮阶段时,大部分藻类是以共生状态存在的,叶状体中的共生藻是光合生物量的主要贡献者.
具鞘微鞘藻(M.vaginatus)在所有结皮类型中都是自由生长藻类群落中的第一优势种,其生物量(以结皮计,下同)最大可达到0.33mm3·g-1,从藻结皮到藓结皮逐渐减少(P<0.05).爪哇伪枝藻(S.javanicum)在结皮中的生物量仅次于微鞘藻,为第二优势种.伪枝藻(S.javanicum)在藻结皮和藓结皮中的生物量均较低,在地衣结皮中的生物量较高,最大可达到0.13mm3·g-1.念珠藻(其中一种:Nostocsp.)在藻结皮、地衣结皮以及藓结皮中的生物量都很低,大多情况其生物量都<0.02mm3·g-1,但在地衣结皮中念珠藻(Nostocsp.)生物量却明显高于其他两种演替类型(P<0.05),最高可达到0.06mm3·g-1(图3).

2.3结皮中异养微生物生物量变化
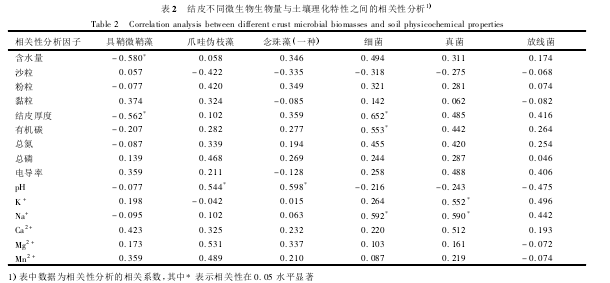
3、种异养微生物在不同演替阶段结皮中的生物
量呈现相似的变化规律(图4),即从流沙到藻结皮,3种异养微生物的生物量都出现一次增加,之后到地衣结皮,它们的生物量又呈现下降的趋势,最后演替到藓结皮,3种异养微生物的生物量再次增加,而且它们的生物量均在藓结皮阶段达到最大值,细菌(以菌落/结皮计,下同)达到18×105CFU·g-1,真菌达到3.4×105CFU·g-1,放线菌达到16.5×105CFU·g-1.
2.4结皮微生物生物量与土壤理化特性之间的关系
随着结皮的发育演替,结皮厚度、含水量逐渐增加,结皮质地中沙粒含量逐渐减少,粉粒、黏粒含量增加(表1).同时结皮中有机碳、总氮、总磷含量也逐渐增加,各种离子在结皮土壤积累(数据未显示).通过结皮微生物生物量与土壤理化指标相关分析发现,微鞘藻(M.vaginatus)生物量与结皮厚度及含水量呈显著负相关(P<0.05);伪枝藻(S.javanicum)、念珠藻(Nostocsp.)生物量都与pH呈显著正相关(P<0.05);细菌数量与结皮厚度、有机碳及Na+含量呈显著正相关(P<0.05);真菌数量与K+、Na+含量呈显著正相关(P<0.05;表2).
3、讨论
在干旱、半干旱荒漠地区,由于水分的缺乏大部分植被很难生存,而生物结皮由于其独特的生理生态特性却可以在该地区广泛分布.在生物结皮发育演替的过程中,很多研究已证实蓝藻(尤其是丝状蓝藻)由于其对荒漠环境极强的适应性能够优先在沙土表面拓殖生长,并最先形成藻结皮.藻结皮代表结皮发育演替的初级阶段,但它们却能够通过改善表土微环境来增加晚期结皮物种拓殖生存的可能性,从而促进结皮的发育演替.在本实验中发现,沙坡头地区的生物结皮一般按照“藻结皮?→地衣结皮?→藓结皮”的模式发育演替,但在一些微环境条件下,藓类也可直接在藻结皮上萌发生长,从而从藻结皮直接演替到藓结皮.
结皮的发育演替受很多周围微环境的影响(如土壤结构、类型,辐射强度,地形等),甚至由于一些资源条件的限制(如水分、营养等),会使结皮停留在某一发育演替阶段.在沙坡头地区,本实验研究发现总体上生物结皮在从早期的藻结皮向晚期的地衣结皮、藓结皮发育演替,而且随着结皮的发育演替,土壤质地逐渐改善,结皮表面特征也相应发生变化(如颜色加深、表面更加粗糙).这些变化一方面改善了结皮区域的土壤微环境,另一方面由于土壤微环境(如土壤营养、理化性质等)的改善进一步促进了结皮的发育演替.尽管整体上研究区域的生物结皮在不断向晚期阶段发育演替,但也有研究表明一些不利的因素仍可能致使晚期的结皮逆向演替到早期阶段(如干扰,长期的干旱、高温等).因此,即使在同一研究区域,由于微环境的差异仍可能导致不同的结皮发育演替模式,从而在同一区域呈现多种不同发育演替阶段的生物结皮.
随着结皮的发育演替光合生物量逐渐增加,因此结皮光合生物量被认为是评价结皮发育水平的一个良好指标.在结皮光合生物中,藻类、地衣、藓类在不同的结皮演替阶段成为结皮的优势类群.在演替初期的藻结皮中,地衣、藓类的生物量较低,主要的光合生物为具鞘微鞘藻(M.vaginatus)和爪哇伪枝藻(S.javanicum);但当结皮演替到晚期时,地衣、藓类逐渐成为结皮的优势类群,如地衣结皮中的坚韧胶衣(C.tenax)以及藓结皮中的银叶真藓(B.argenteum)和黑对齿藓(D.nigrescens).此时,地衣或藓类生物量显著超过藻类,占据结皮大部分光合生物量,但如果将地衣叶状体和藓类“茎叶”部分去除后,自由生长的藻类便是结皮中仅有的光合生物类群.本实验研究发现地衣结皮中(地衣盖度约70%~80%)自由生长的藻类生物量不到光合生物量的30%;藓结皮中(藓类盖度100%)藻类生物量还不到光合生物量的10%.
地衣是藻类与真菌形成的共生体,因此就藻类生物量而言,尽管本研究发现自由生长的藻类生物量从藻结皮到地衣结皮有所下降,但总的藻类生物量却明显增加.这就表明在地衣结皮中,大部分藻类是以共生状态存在的,地衣叶状体中的共生藻是光合生物量的主要贡献者.之后,随着结皮进一步向藓结皮发育演替,实验结果显示结皮中的藻类生物量呈下降的趋势,这可能是由于随着藓类的出现,藻类的生存空间逐渐被占据所致.然而由于藓类的出现,结皮厚度、含水量明显增加(P<0.05),因此本实验中微鞘藻(M.vaginatus)生物量显示与结皮厚度及含水量呈负相关关系.另外,本实验中微鞘藻(M.vaginatus)生物量随着结皮的发育演替逐渐下降,这也表明微鞘藻(M.vaginatus)胶结、捆绑土壤颗粒的作用逐渐被地衣、藓类所取代.伪枝藻(S.javanicum)和念珠藻(Nostocsp.)为固氮种类,它们在晚期地衣结皮中生物量的增加则可能是由于结皮发育演替对氮需求的增加所致.
随着结皮的发育演替,表土微环境被逐渐改善,而各种土壤理化特性(如有机质含量、土壤质地等)又是影响土壤微生物生物量的重要因素.本实验研究发现异养微生物生物量(细菌、真菌、放线菌菌落数)随着结皮的发育呈现先增加后减少,最后再次增加的趋势,而且在不同演替阶段结皮中的关系为:藓结皮>藻结皮>地衣结皮.与藻结皮相比,异养微生物生物量在地衣结皮中明显下降,这可能是由于大量地衣的出现所致.因为有研究报道地衣中大量的假根(共生真菌)不容易被培养,而这些假根也有可能通过分泌化学物质来抑制其他异养微生物的生长.实验中细菌、真菌以及放线菌的数量都与总的异养微生物生物量呈一致的变化规律,其中细菌数量的增加与有机碳、Na+含量的增加有关,真菌数量的增加与K+、Na+含量的增加有关(P<0.05).
4、结论
(1)通过对腾格里沙漠东南缘沙坡头植被恢复区生物结皮的调查发现,该地区的生物结皮按照一定的模式进行发育演替,从早期到晚期有藻结皮、地衣结皮和藓结皮3个主要的结皮发育演替阶段.
(2)随着结皮的发育演替,结皮光合生物生物量逐渐增加.当藻结皮演替为地衣结皮时,结皮中虽然藻类生物量(包括地衣中共生的藻类生物量)增加,但自由生长的藻类(不包括地衣中共生的藻类)生物量却下降,此时大部分藻类处于共生状态;当藓类大量出现后,藻类的生存空间被占据,因此藓结皮中藻类生物量进一步下降.
(3)随着结皮的发育演替,土壤微环境被逐渐改善,为异养微生物的生长提供了良好条件.结皮中不同微生物类群生物量的变化一方面是对土壤微环境变化的响应;另一方面也体现了结皮不同发育演替阶段生态功能的需求,这对结皮在荒漠地区的适应性生存有着重要的意义.





