两性异形在脊椎动物中普遍存在( Darwin1871,Anderson et al. 1990,Andersson 1994) ,蜥蜴类动物中亦存在两性间个体大小、局部形态特征和体色上的差异( Martin et al. 2009,蔡垚 2009,Faizi et al. 2010,Qu et al. 2011) 。两性谋求各自的繁殖利益是两性异形产生的重要原因之一。例如,较大的雄性在同性的资源竞争( 配偶、食物、优质生境等) 中具有优势,这也促进了雄体大于雌体的两性异形的形成( Shine1979,Katsikaros et al. 1997,Gvozdík et al.2003,Gienger et al. 2007 ) 。 生 育 力 选选择( Woolbright 1983,Olsson et al. 2002) 、寿命或死亡 率 ( Dunham 1981) 、生 长 速 率 ( Brooks1991,杜卫国等 2001) 及食物生态位( Berry etal. 1980,Powell et al. 1985,林植华等 2000)等方面的差异也有可能导致两性异形。
关于沙蜥类两性异形的研究主要有奇台沙蜥 ( Phrynocephalus grumgrzimailoi) ( 刘 洋 等2009) 、荒漠沙蜥 ( P. przewalskii) ( Zhao et al.2013) 、青海沙蜥 ( P. vlangalii) ( Zhang et al.2005) 、变色沙蜥( P. versicolor) 和无斑沙蜥( P.immaculatus) ( 屈 彦 福 等 2005, Qu et al.2011) 。而关于草原沙蜥( P. frontalis) 的研究报道,仅限于简单的食性( 赵雪等 2013) 、活动节律( 连雪 2011) 等方面。草原沙蜥两性异形的研究仅屈彦福等( 2005) 、Qu 等( 2011) 有过相关报道。本文测定了草原沙蜥幼体的部分形态学数据,对其个体发育过程中幼体的两性异形进行探讨。
1 材料与方法
1. 1 实验材料 研究用的 118 条草原沙蜥幼体( 79♂ ,39♀) 于 2011 年 8 月采自内蒙古鄂尔多 斯 北 部 的 库 布 齐 沙 漠 ( 40° 12' N,111°06'E) 。捕捉的蜥蜴带回内蒙古师范大学动物学实验室并鉴定每条蜥蜴的性别。
1. 2 测定方法 用游标卡尺( 哈尔滨量具刃具有 限 公 司,量 程 0 ~ 150 mm,精 确 到0. 02 mm) 测量蜥蜴的体长 ( snout-vent length,SVL,即吻端至泄殖腔孔前缘间距 ) 、头宽( head width,HW,左右颌关节间距) 、尾长( taillength,TL,泄殖孔至尾端部的距离) 和腋胯距( abdomen length,AL,前肢后缘基部到后肢前缘基部之间的直线距离) 。草原沙蜥成体的最小体长为43. 5 mm ( Qu et al. 2011) ,故以此为界区别成体和幼体。
所有数据用 Excel 软件和 Spss 软件处理,数据作参数统计分析前,分别检验其正态性( Kolmogorov-Smirnov text) 和方差同质性 ( F-max text) 。用 t-检验、线性回归、协方差分析( ANCOVA) 处理相应的数据,描述性统计值均用平均值 ± 标准误表示,显着性水平设置为 α= 0. 05。
2 结 果
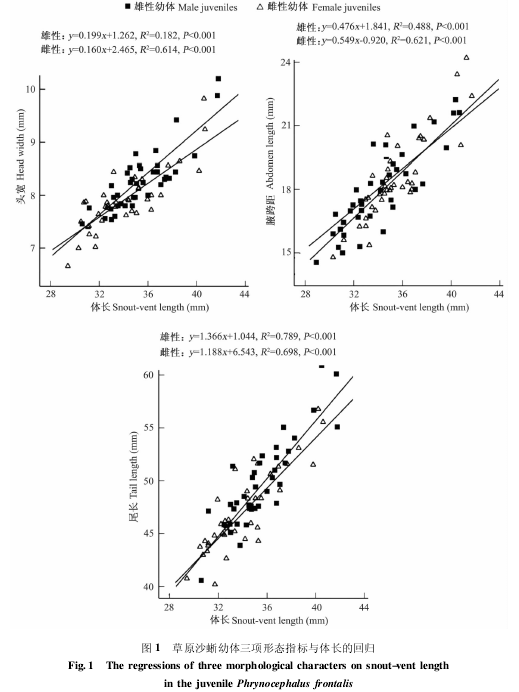
雌雄两性草原沙蜥幼体的平均体长( SVL)存在显着的两性差异( t =3. 702,df =116,P <0. 001) ,雄性幼体的体长显着大于雌性幼体( 表 1) 。草原沙蜥幼体的局部形态特征皆与体长成正相关。以体长为协变量的 ANCOVA 显示: 雄性幼体头宽( F1,115= 357. 71,P < 0. 001)及尾长( F1,115= 372. 74,P < 0. 001 ) 随体长的增长速率大于雌性幼体,但其腋胯距( F1,115=160. 93,P < 0. 001) 随体长增长的速率小于雌性( 图 1) 。由于两性幼体的头部随体长的增长均呈异速增长,故 ANCOVA 不适用于比较特定体长的两性头宽及腋胯距等指标。t-检验结果显示,雄性幼体的头宽( t =4. 616,df =116,P =0. 002) 及尾长 ( t =3. 958,df =116,P <0. 001)均显着大于雌性幼体,而其腋胯距( t =2. 292,df = 116,P = 0. 024 ) 显着小于雌性幼体 ( 表1) 。
3 讨 论
蜥蜴的成体体型大小分为 3 种类型: 雄性成体大于雌性成体,雌性成体大于雄性成体,两性成体无显着差异( Powell et al. 1985) 。草原沙蜥成体属于第一种类型( 屈彦福等 2005) ,与地中海壁虎( Hmidactylus turcicus) ( Daniel1996) 、普通岩蜥 ( Podarcis muralis) ( Bra?a etal. 2000) 、变色树蜥( Calotes versicolor) ( 邱清波等 2001) 、中国石龙子( Eumeces chinensis)( 林植华等 2000,计翔等 2002) 及蓝尾石龙子( E. elegans) ( 杜卫国等 2001) 相同。但 Qu 等( 2011) 的研究结果中则表明,草原沙蜥的成体体型属于第三种类型,两种研究结果的不同可能与研究地的生境、食物类型及繁殖压力不同有关。在研究过的沙蜥中属于第二种类型的只有青海沙蜥,而无斑沙蜥、变色沙蜥、奇台沙蜥则都属于第三种类型。
雄性具有较大的体形与其在食物竞争、雄-雄性内竞争或者通过雌性选择获得更大的交配成功 率 有 关 ( Andersson 1994,Olsson et al.2002) ,体型较大的雄性往往会在竞争中获得胜利,最终导致其繁殖成功率大于体型较小的雄性。而本研究结果显示,草原沙蜥的两性异形从幼体时就开始出现,表现为雄性幼体具有较大的体型,但由于没有初生个体的数据,故其两性异形的起点尚不清楚。
蜥蜴中雄性头部普遍比雌性蜥蜴大( 计翔等 2002,Verwaijen et al. 2002,Zhang et al.2005) 。蜥蜴头部大小在一定程度上可视为资源持有能力的标志,头部较大的雄性通常被头部较小的雄性视为竞争中潜在的胜利者,能够优先获得包括配偶在内的多种资源 ( 计翔1994) 。已有研究表明,头部较大的蜥蜴其张口和 咬 合 力 较 强 ( Verwaijen et al. 2002,Gvozdík et al. 2003,Dubey et al. 2011) ,这使得头部大小不同的蜥蜴在食性上必然会产生分化,而具有较大头部的蜥蜴也必然具有较强的资源竞争力。草原沙蜥头部的两性异形在幼体时 就 已 经 出 现, 这 一 特 点 与 蝘 蜓( Sphenomorphus indicus) ( 计翔等 2000) 、中国石龙子( 林植华等 2000) 等相似。雄性在幼体时即有较大的头部更利于其扩大食谱、增强获得较多食物的能力,以满足其性成熟后拥有更大竞争力的需要。而雌性为了获得最大的繁殖利益,在幼体时就已减缓头部的生长来确保腹部的生长,以在性成熟时进一步增大腹腔容纳量。雌性具有较大的腹长可能与生育力选择有关,因为雌体的腹腔容纳量能限制其繁殖输出( Du et al. 2010) 。本研究结果显示,雌性草原沙蜥幼体具有较大的腋胯距和较小的头部,这有可能直接有利于其增大繁殖输出,而这也可能是导致两性幼体头部和腋胯距异形的主要原因。
尾部与爬行动物的运动能力相关( Arnoldet al. 1988) 。有研究表明,雄性沙蜥具有较长的尾部可能与容纳半阴茎、牵缩肌有关( King1989) 。性选择也可能对尾长产生影响,雄性用尾部向雌性示爱,一个强健的尾部可以提高这种能力( Gillingham 1987) 。在本研究结果中,雄性幼体具有较长的尾部,尾长的回归线斜率也大于雌性幼体。这表明雄性在生长过程中同样分配部分能量用于尾部的增长,使其在成年后更易获得雌性的青睐,以提高其繁殖成功率。
综上所述,草原沙蜥幼体具有显着的两性异形,这一现象也较为普遍( 张永普等 2000,许雪峰等 2003) 。雄性幼体具有较大的头部、尾部及较小的腋胯距,说明雄性在生长过程中将一些能量分配于体长、头部的增长,而雌性则从幼体时就已将能量分配至一些对繁殖较为重要的部位,如腋胯距,以实现在成年后获得潜在较大的窝卵数或窝卵重。这表明两性在生长过程中会将能量分配于不同的部位,以使各自在成年后获得更大的利益。
参 考 文 献
Andersson M. 1994. Sexual Selection. Princeton,New Jersey:Princeton University Press.
Anderson R A,Vitt L J. 1990. Sexual selection versus alternativecauses of sexual dimorphism in teiid lizards. Oecologia,84( 2) : 145 -157.