��Ⱥ��̬ѧ�����о��ȵ�10ƪ֮����ƪ���ҹ��غ��ֲ��Ĵ���������Ⱥ��������
����ժҪ��������Ⱥ��̬ѧ��� ���ҹ��غ��ֲ��Ĵ���������Ⱥ���ֽ��������������Ƿ��е���������Ϊ���ֱ�Ҫ������ ������Ϊ�����ο�����������֤��Ϊ�������ֲ��������Ȼ���ϵĵ������벢�����ڡ��ƺ���������̨��ů�����ذ����γɵĻ����� ̨�庣Ͽ���Ϻ��ɼ����γɵĻ����� ���ҹ��غ��γ�����ȫ�ָ��ĺ�����ϵ�� ��Ϊ�����γɵ������Եĵ������ϣ� ���ϱ����������γ��ϻƺ�-�����Լ�̨�庣Ͽ-�Ϻ�2��������Ⱥ���ϻƺ�-����������Ⱥ�ɳ������ϲ����й��������������Ŀ��èͷ��ͷ�پ���Ͷ����е���8������Ⱥ����ɣ�̨�庣Ͽ-�Ϻ�������Ⱥ��ţɽ��������������졢�ϰĵ�����β�⺣�����ݵ��������������ź�����6������Ⱥ����ɡ�
�����ؼ���������㣻 ��Ⱥ���֣� ��Ⱥ��̬ѧ��
����Abstract:
����The geographic population dividing for large yellow croaker ��Pseudosciaena crocea�� is discussed from the perspective of population ecology.From the concept of "geographic population" in population ecology, the geographic isolation is the necessary condition for dividing the population into the different geographic groups with other factors used as the auxiliary reference.Although the solutions are not absolute, the area of P.crocea is divided into two completely separate oceanic current systems: ��1�� the ocean circulation formed by Taiwan warm current and coastal current in the East China Sea and Yellow Sea, ��2�� the ocean circulation formed by South China Sea monsoon in the Taiwan Strait and the South China Sea.The two oceanic current systems separate P.crocea into two geographic populations: southern Yellow Sea and East China Sea ��SYS-ECS�� population, and Taiwan Strait and South China Sea ��TS-SCS�� population.The SYS-ECS population includes 8 spawning aggregations as follows: south-west area of Korean Peninsula, Lvsiyang, Daiquyang, Damuyang, Maotouyang, Dongtouyan, Guanjingyang and Dongyin archipelago.In contrast, the TS-SCS population covers 6 spawning aggregations: Niushan Island, outside islands of Jiulong River, Nanao Island, Shanwei-offshore, Naozhou Island and Xuwen-offshore.
����Keyword:
����Pseudosciaena crocea;division of populations;population ecology;

��������� ��Pseudosciaena crocea�� , ��ʯ����ơ��������� �׳ƻ��㡢�ƻ���ȣ� Ϊ�ҹ�����ůˮ�Լ�Ⱥ�̾�����ε����࣬ �����ڽ������С��²����������ҹ������ĺ����Ĵ��̶���֮һ�� 20����70���ǰ������������������19���[1]���ڴ��������ҵ�е���Ҫ��λ�� ����50������Ϳ�ʼ�Դ������Դ�����˳������飬 �ڲ�ͬ�Ρ����ݲ�ͬ�����ݣ� �Դ�������Ⱥ�����˻��ֵ�����ѧ�������Ⱥ�Ļ������ݺ���Ⱥ�Ļ�����δͳһ��20����50����� �й���ѧԺ���������й�����㻮��Ϊ2����Ⱥ�����ƺ�Ⱥ������Ⱥ[2].60����������β���������ҹ�������Ϊ3���������壺����塢��-�������������[3,4].�����β����Ľ���ܶ������ص�Ӱ�죬 ������̬ѧ��Ⱥ���в�ȷ���ԡ����ݱ�־�����ز������·�ߣ� ��������-������ӦΪͬһ����-�ƺ���Ⱥ�� ���ɷ�Ϊ2����ͬ�IJ���Ⱥ��[5]��1������������[6]�ο��ֲ���Χ�����·�ߺ���Ե��ϵ�� ������㻮��Ϊ�ϻƺ�-������̨�庣Ͽ-����������3��������Ⱥ
�������Ŵ��������ֳҵ�ķ�չ�� ����������Դ���ӣ� ����ֳ������������Դ�ֱ治�壬 ֱ��Ӱ���˴���������ѡ������ֳ�����������ߵ�������������ֳ������������Դ�ĸ���Ӱ�졣��ˣ� �Դ������Ⱥ��Ⱥ����п�ѧ�����ѿ̲��ݻ������ߴ���Ⱥ��̬ѧ������������� �Դ���������Ⱥ�������Ⱥ��Ļ��ּ��������� ּ��Ϊ����ѡ������ֳ��������Դ���������ṩ��ѧ���ݡ�
����1 ������̬ѧ����Ⱥ��Ⱥ��Ļ�������
����1.1 ��Ⱥ�Ļ�������
������̬ѧ��"��Ⱥ"�Ļ��֣� ���������ֲ��Ƿ������Ϊ�ж���Ⱥ�Ƿ�������ڵ�����Ҫ��[5].��Ⱥ����һ���ռ���ͬ�ָ���ļ���[7].������[8]��Ϊ�������Ȼ��ȺӦ����3�������� ��1�� �ռ���������Ⱥռ��һ���ķֲ����� ����ֲ����ޣ� ��������Ⱥ�������Ǹ���ġ� ��2�� ���������� ����Ⱥ����ʱ����һ�µ��������࣬ �����һ���������䶯���ɡ� ��3�� �Ŵ������� ����Ⱥ�����ڷ�ֳ��Ԫ�� ͬ����һ����ʱ���������Ļ���⡣
����1.2 Ⱥ��Ļ�������
��������ҵ����ѧ��ʹ�ø����ձ����"Ⱥ��"�����������"�ܹ�����һ����ҵ����ģʽ���Dz����㣬 �ɶ���Ϊһ��Ⱥ�塣Ⱥ������Ⱥ֮�µ�һ���о���λ"[9].����Ⱥ����Ϊǿ����ҵ�������������������һ��ˮ����Դ�о���λ�� ������ҵ��Դ���������������о���ʵ�����γɵĻ������
����1.3 ��Ⱥ��Ⱥ�������
�������¾�[9]��Ϊ�� ��Ⱥ������̬ѧ����ĸ�� ���л�����Ⱥ��֮���һ��������Ρ�Ⱥ����ƫ������ҵ�����ĵ�Ԫ�� ���ܹ�����һ����ҵ����ģʽ���Dz����㣬 ������Դ��������ҵ�����о���ʵ�����γɵģ� �ܵ��������ú�����Ӱ�졣��������Ⱥ��Ⱥ��֮�䣬 ��Ⱥ�ǿ۵�����ѧ��Ԫ�� Ⱥ������ҵ������Ԫ�� ���߹�ϵ���У� ���������ڴ�����ϵ����ijһ��ֳȺ�壬 ����������Ⱥ֮�µ�һ������ѧ��ֳ��λ������ijһ����Ⱥ��Ϳ�������������е�"Ⱥ��"��ȿ�������Ⱥ֮�µ�һ��Ⱥ�壬 Ҳ������һ����Ⱥ��
������ˣ� �ڻ��ֵ�λȺ��ʱ�� ��Ҫ������̬����������̬���Ŵ��������ԣ� ��Ҫ��֤�ؿ�����ҵ������ʵ����Ҫ�� ȷ�����ʵ��ĵ�λȺ�塣
����2 �����ĵ����ֲ����������
����2.1 �����ĵ����ֲ�
�����й������ķֲ��� ���㽭�غ�Ϊ���� ��Ҫ�泡�ں��������ϲ��Լ��й��Ľ����������㽭�����Ŀ��èͷ��ͷ������Ķ����е����پ���ţɽ��������������� �����Ÿ������죩 ���泡���㶫���ϰĵ�����β���������ݵ��������������ź������泡[10].
����2.2 ��������Ȼ���ϵĵ�������
�����Ӵ����ĵ����ֲ����� ��Ȼ���ϵĵ������벢�����ԡ�̨�嵺�ͺ��ϵ�2��������һ���ķָ����ã� ���û������ԭ����Էָ��� ������Կ�ͨ��̨�庣Ͽ������Ρ��ͺ��ϵ����ԣ� �����ֲ��ĺ��������ں��ϵ����� ���ں��ϵ�����������Ȼ���ϵĵ������벢������Ϊ��Ⱥ����Ҫ���ݡ�
����2.3 ������ɺ����γɵ���������ָ�
�����й��������ڽ������Ļ����ֲ�[11], ������ֲ��йء�
����2.3.1 �ơ������Ļ����ֲ�
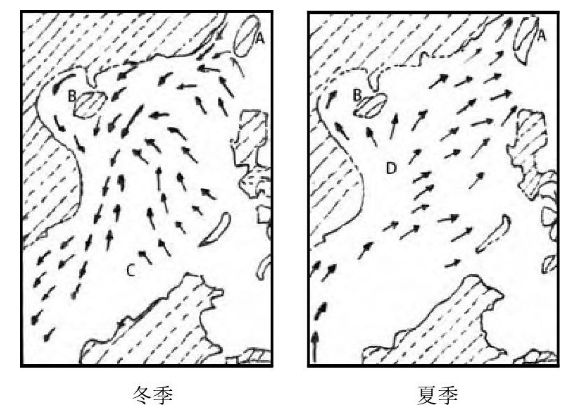
�����ơ������ĺ�����Ҫ���ɺڳ�ů�� ��ͼ1B�� ���ذ���2����ϵ��ɣ� ��������ʽ�������������ڳ��������Ĺ����У� ����һ���������ʿ��Ͽ�⣬ ��������̨�嶫�������� ������붫���� ���ش�½������ ��ͼ1�� .�����ձ�������֮�������ֿ�ʼ��֧�� ����һ֧�ڼ��ݵ����汱�Ͻ���ƺ��ϲ��� ��Ϊ�ƺ�ů�����ƺ�ů���ڱ��Ϲ������ܵ��ذ�ˮ�ļ�����������Ӱ������ԣ� ů���������Ž���ƺ��ľ������Ӷ��ܿ������

����ͼ1 �������ƺ��Ͷ����ĺ���ϵͳʾ��ͼ
�����ڳ����� �������� ���ҹ�̨�� ��ͼ1E�� ��������ֳ�һС��֧�� �������⺣���ϣ� �ɴﺼ�����⣬ Ȼ��ת������ƺ���ˮ��϶����ԣ� ��֧������̨�帽�������� ��Ϊ̨��ů���� Ҳ��Ϊ�ڳ��������֧����֧ů�����ҹ��㽭�����������¡����ε��⺣ˮ���������ذ�������ʱ�� ���������Եķ��������dz�Ϊ"����", �γ���������ɽ�泡���ڳ����ҹ�̨��Ķ��Ϻ��� ��10��������4�£� ��һ����������ʿ��Ͽ�����Ϻ��� Լ��̨��������ַ�Ϊ2֧�����������ϣ� ��Ϊ�Ϻ�����������һ����ɲ��֣�֧����̨���������ϣ� ��ڳ����ɺ�̨��ů����ϡ��ڳ��ܵ����ε�Ӱ�죬 ���ص����������ٺ�����������ͬ��
���������ҹ��ذ��������С��ͬ�Ľ����뺣�� ���ذ���ˮ�嵭�� ��Щ���嵭�ĺ�ˮ�ذ�����ȥ��Ϊ�������⺣ů���Ľ�����ƽ�⣬ ���˶������������ɱ����ϣ� �����ϵ����⺣��ˮ��ϣ� ��������С���С����ҹ��غ��Ա������������йص���Ҫ���ձ��ذ��� ��ͼ1C�� �������ذ��� ��ͼ1D�� �ȡ�
�����ձ��ذ���Դ�ں����塢���ӿ�һ���� �ϽӲ����ذ����� �غ�������30 m���������£� �������ں� ������ȥ�� ��ڳ����Ϸ�֧���Ϻ�����ʧ���ձ��ذ���ÿ��6~8�£� �ܳ���������Ӱ�����ǿ�� ��Ӱ��ɴ��������ʺ�Ͽ ��ͼ1A�� .
���������ذ�����Դ�ڳ����ںͺ�����һ���� ��Ҫ�ɳ�����Ǯ�����ľ����뺣��ɣ� ��;����걽��������ĵ�ˮ���룬 �����ذ���������ƽ̶����̨�嵺������������Ϻ��� ���෴�����̨�庣Ͽ���Լ��ڳ���֧��̨��ů�������� �谭�������ذ������������ƽ�[12].���ֲ��ڳ����ڼ������ϵ������ذ����������ذ������ڶࡢ�������ۼ������¶Ƚ϶��� �������Ŀ��Ƚ�խ�� һ��శ37~74 km.
����2.3.2 �Ϻ��Ļ����ֲ�
�����Ϻ�λ���ȴ��������� �ļ�ʢ�����Ϸ磬 ����ʢ�ж����磬 ���緽���뺣������һ�£� �������ȶ���ϵ�ķ�չ���Ϻ����滷���ڷ�������£� ���м���Ư�������� ��ͼ2, AΪ̨�壬 BΪ���ϣ� .�ڼ���ij��ڴ����£� �Ϻ���������·����ǿ�ȶ��漾����䡣���ϼ����ڼ䣬 �Ϻ�Ϊ������ ��ͼ2D�� , ���������ڼ䣬 �������Ϊ������ ��ͼ2C�� , �ڳ����ֺ�ˮ����ʿ��Ͽ�����Ϻ������� ͬ����̨�庣Ͽ���ذ����ϲ��������ϡ��ӻơ��������Ϻ��Ļ����ֲ����� ���ֲ��ڽ����Ĵ���㣬 ��̨�山��Ϊ��ָ�Ϊ�ϱ���Ⱥ��
����ͼ2 �Ϻ����绷��ʾ��ͼ
����3 ����㲻ͬȺ���ĸ��彻�����Ŵ�����
����3.1 ��ͬȺ���ĸ��彻��
����1959��4��21�ո���ʡˮ���о����ڸ���ʡ�����ر����ز����㽭ʡ����ˮ���о�����1958��5��20���������������ѳ���־�������Գ�������Դ���㣬 ��β��1958����������������ֳ�� ʱ����һ���ʱ�����ε��������ѳ�������ֳ�� ֤ʵ��������������Ⱥ�����������ڵĹپ������Ⱥ������������嶫�����Ķ����е�����Ⱥ��������ֳ�����ϵ�� ��2�� .ë���ֵ� ��1�� ��־��������ʵҲ֤ʵ����һ�㡣
������һ�ز���¼˵�����������پ�������Ⱥ�����������������������������[5]�о��˶��ƺ���������·�ߣ� ����10�����ҵ��˾1971~1982�����㲶��ͳ�����ϣ� �Ӵ��������ֲ�����Ⱥ�ƶ��ȷ������������ ���ƺ������ֻ��1��������Ⱥ��2��Խ�������⺣Խ������Ҫλ��30°00′~32°00′N, 124°00′~126°00′E������Խ����λ���㽭���ϲ������������������ÿ��3~4�£� �⺣Խ����Ⱥ������������ɽ�泡�ͳ������泡��5�²���Ⱥ�����������������泡���ѣ� ��һ����Ⱥ������������ɽȺ���غ����������������ʹ�Ŀ��Ⱥ�����ѡ�6���ڳ����ں������泡������6~8������Ⱥ�山�ϻƺ��ϲ�����������10���Ժ� ������������£� �����Ĵ����������Խ����Σ� ����Ⱥ�������⺣Խ������ ��һ����Ⱥ��������»ص��������ϲ�������Խ�������㽭���ϲ������������������Խ���Ĵ����Ⱥ�壬 �ڴ�����������У� ����Ⱥ�山������ɽ�泡���⺣����Ⱥ���ϣ� ���������泡����ɽȺ���غ����ѣ� ����Ⱥ��ͽ�����èͷ��ͷ�������塢�پ���Ͷ������Ⱥ�����ѣ� ���Ѻ��Ⱥ���ڲ��ѳ����������� �����ֻص��ͽ���Խ�����������·�߿��� ̨���Ա������IJ�ͬȺ�����������ܺ���ֳ�����������
����3.2 ��ͬȺ�����Ŵ�����
����Ŀǰ���ж�����پ���ͬ�����Ⱥ��ʹ��RAPD��AFLP��SSR��ISSR��COI�ȷ��������Ŵ������о��ı���[13,14,15,16,17,18,19,20], ���������ʾ��ЩȺ�����Ŵ�����ܽ��� ���첻������������������������Ⱥ�� Ŀǰ��ȱ���й����ϡ����磺�����ĵ�[18]��SSR������ ���������ṩ�Ĵ����4��Ⱥ������Ŵ���������� �����ʾȺ����Ŵ�����ϵ�����ϸߣ� ��������֮��IJ����С������Զ��[19]������Ⱥ�����Ⱥ����㼰���ӽ��Ӵ������������Ŵ���������� �����ʾ����Ⱥ�ͺ��Ⱥ���������ϵͣ� UPGMA������ʾ���Ⱥ������Ⱥ�Ŵ���������� ��ʾ��ֳ�����Ⱥ������Ⱥ�������Ŵ���������ϵ͡���������[20]����������COI��������Ƭ����ISSR��Ƿ����������泡 ���ƺ�Ұ���� ������� ����ֳ�� �پ��� ����ֳ�� �������Ŵ������Խ����о��� �����ʾȺ�����Ŵ��ֻ�ˮƽ�ܵ͡������Ϻ��Ĵ���㣬 Ŀǰ��ȱ���ⷽ����й����ϡ�
����4 ����������Ⱥ���ּ���Ⱥ��Ļ���
����4.1 ����������Ⱥ�Ļ���
�������ϣ� �Ե����ֲ��Ƿ���ڸ�����Ϊ�ж���Ⱥ�Ƿ�������ڵ�һ����Ҫ������ ������Ϊ�����ο��� ����ȫ������Ⱥ��̬ѧ���ڻ��ֵ�����Ⱥ�ĸ���������ķֲ����������̨�庣Ͽ����Ϊ���̨��ů�����ذ���֮���γɻ�����̨�庣Ͽ-�Ϻ��γɵļ��绷���� �γɵ�����������룬 ���ֲ��ϱ��������뿪���� ��Ӧ���γ�2��������Ⱥ�� ���ϻƺ�-����������Ⱥ��̨�庣Ͽ-�Ϻ�������Ⱥ��
������̨�庣Ͽ�����Ĵ���㻮��Ϊ�ϻƺ�-����������Ⱥ�� �����й���ѧԺ�����о���[2]���������[5]����������[6]�Ļ�����һ�¡�20����60��������ϵ�[3]���칧�ѵ�[4]������̬ѧ������̨�庣Ͽ�����Ĵ���㻮��Ϊ��������-������Ĺپ����붫����Ⱥ�壬 ����̬������״���������ص��ģ� �����β����Ľ���ܵ���������Ӱ�죬 ������Ϊ��Ⱥ�����ݣ� ����֮�䲢�����ں���ˮ�Ķ���ѧ�ĵ������롣����־����[13]���������ι���[5]���Ŵ�����[13,14,15,16,17,18,19,20]����֤�� Ҳ����֧�ֹپ�����ɽ�������������ͬ�ĵ�����Ⱥ�����پ�������������ϻƺ�-����������Ⱥ�ǿ�ѧ�ġ���������[6]��Ϊ��������ͳ���̾������˽�Ĵ�������·�ߣ� �����е��پ����������Ⱥ���Ӧ�������ϻƺ�-����������Ⱥ��
����̨�庣Ͽ-�Ϻ�������Ⱥ�DZ����״�����Ļ��ֹ۵㡣20����50����� �й���ѧԺ���������ݵ�ʱ�о����ֵ��������ֲ��д���㣬 ������֮Ϊ����Ⱥ����̨�庣Ͽ�������������ϵ����д����ֲ��� ��������[6]��̨�庣Ͽ-�Ϻ��ֲ��Ĵ���㻮��Ϊ2��������Ⱥ�� ��̨�庣Ͽ-����������Ⱥ������������Ⱥ�� ��������Ϊ��̨�庣Ͽ-����������Ⱥ�ɴ������������Ĵ��ں�����Ⱥ����ɣ� ������������Ⱥ���^��ֳ������Ⱥ��Ϊ���� ����Ⱥ��Ϊ���� �ɴ˶���֮������û�к���ˮ�Ķ����γɵ�����������룬 ��֮���д������ﷱ��Ⱥ�壬 ��ֳ���벢�����ԣ� ���ܺ���ֳ�����Dz��ɱ���ģ� ���Ա�����Ϊ��2��������Ⱥ��Ϊһ��������Ⱥͳ��Ϊ"̨�庣Ͽ-�Ϻ�������Ⱥ"��Ϊ����ʵ�ʡ�
���������ϻƺ�-����������Ⱥ��̨�庣Ͽ-�Ϻ�������Ⱥ2��������Ⱥ�����̬��������̬�ͷ�ֳϰ�ԣ� �Լ��Ŵ��Ȳ����д���һ���о���
����4.2 �ϻƺ�-����������Ⱥ��Ⱥ�����
���������й�ѧ���о�ȷ���IJ��ѳ�[3,4,6,21], ����ʷ�γɵ��泡�� ��Ϊһ��Ⱥ��ķֲ����� �ϻƺ�-����������Ⱥ�ɳ������ϲ�����Ⱥ�塢�й������������Ⱥ�塢��������Ⱥ�塢��Ŀ�����Ⱥ�塢èͷ�����Ⱥ�塢��ͷ�����Ⱥ�塢�پ������Ⱥ��Ͷ����е�����Ⱥ���8��Ⱥ����ɡ�
����4.3 ̨�庣Ͽ-�Ϻ�������Ⱥ��Ⱥ�����
���������о�����[3,6,22], ̨�庣Ͽ-�Ϻ�������Ⱥ�Ĵ������ţɽ������Ⱥ�塢��������� �����Ÿ������죩 ����Ⱥ�塢�ϰĵ�����Ⱥ�塢��β�⺣����Ⱥ�塢����������������Ⱥ�塢���ź�������Ⱥ���6��Ⱥ����ɡ�
���������
����[1]Liu M, Sadovy de Mitheson Y.Profile of a fisherycollapse:Why mariculture failed to save the large yellowcroaker[J].Fish and Fisheries, 2008, 9:219-242.
����[2]�й���ѧԺ�����о������������������ij����о�[J].��ѧͨ���� 1959, 20:697.
����[3] �����ϣ� �칧�ѣ� �����㡣�������̬�����ĵ�������͵�����Ⱥ����[J].�����ѧ������ 1962 ��2�� :79-97.
����[4] �칧�ѣ� �ޱ����� ���������������Ⱥ�ṹ�ĵ�������[J].�����ѧ������ 1962 ��2�� :98-109.
����[5]������ �¼ѽܡ����ƺ���������·�ߵ��о�[J].ˮ��ѧ���� 2011, 35 ��3�� :429-437.
����[6]�������� �������� ��ʥ�ƣ� �ȡ�����������Ⱥ��̽��[J].�ִ���ҵ��Ϣ�� 2011, 26 ��2�� :3-8.
����[7] ����ʦ����ѧ�� ����ʦ����ѧ�� ������ѧ�� �ȡ�������̬ѧ ���ϲᣩ [M].������������������磬 1980.
����[8]�����ơ�������̬ѧ[M].�������й�ũҵ�����磬 1995.
����[9]���¾�����ҵ��Դ���泡ѧ[M].��������������磬 2004:10-13.
����[10]�Դ��s���й�������ҵ��Դ[M].���ݣ��㽭��ѧ���������磬 1990:40-45.
����[11] ��������ѧУ�ࡣ����ѧ[M].��������������磬 1986:174-178.
����[12]Liu Y G, Chen S L, Li J, et al.Genetik diversity in threeJapanese flounder ��Paralichthys olivaceus�� populationsrevealed by ISSR markers[J].Aquaculture, 2006, 255 ��1/4�� :565-572.
����[13]�����ƣ� �ź����� Ѧ���壬 �ȡ�������ֳ������Ŵ������Ե�ͬ��ø��RAPD����[J].�й�ˮ����ѧ�� 2003, 6:523-525.
����[14]���ij��� �����ƣ� �ܵ����� �ȡ������4����ϵ����̬�������[J].������ѧѧ���������棬 2009, 22 ��2�� :185-190.
����[15]�����ƣ� �����䣬 ������ �ȡ����������پ��������Խ����ӽ��Ӵ��������ܼ��ӽ����Ʒ���[J].ˮ��ѧ���� 2010, 34 ��6�� :859-864.
����[16]��ʫ���� ����Ӣ�� �ź����� �ȡ������ ��Pseudosciaenacrocea�� �����ѡ��Ⱥ��پ�����ֳȺ����Ŵ��������[J].��������ӣ� 2006, 37:41-46.
����[17]�������� л���ܣ� ����ȫ����-���������������ֳ�������Ŵ��������о�[J].���Ŵ�ѧѧ������Ȼ��ѧ�棬 2006, 45:836-840.
����[18]�����ģ� ����÷�� ����Ⱥ�����Ƿ����ĸ������Ⱥ����Ŵ�������[J].ˮ��ѧ��־�� 2009, 22 ��2�� :7-11.
����[19]����Զ�� ����ȫ�� �Ž��裬 �ȡ�����Ⱥ�����Ⱥ��ֳ����� ��Pseudosciaena crocea�� �����ӽ��Ӵ��Ŵ������SSR����[J].��������ӣ� 2011, 42 ��4�� :592-596.
����[20]�������� ��ʿϼ�� ��־�£� �ȡ������Ұ��Ⱥ������ֳȺ���Ŵ��������о�[J].�����ѧ�� 2011, 35 ��12�� :82-87.
����[21] �����꣬ ��۴��� ë���֣� �ȡ�����㶫������ҵ��Դ���������[M].�Ϻ�������ʦ����ѧ�����磬 1987.
����[22]��������տ���غ��������Ⱥ���о�[J].տ�������ѧѧ���� 1998, 18 ��2�� :l5-19.
����ע��
����1 ë���֣� �������� ������������泡�����ʹ������ҵ����ͨ�� 1981.
����2 ����ʡˮ����ѧ�о������پ��������泡���鱨�� ��1957~1959�꣩ .���ţ� 1960.
����3 ��۴��� �±��ܣ� �ų�ï�����������������Ȼ��Դ���鱨�档���ţ� 1985.





