
人乳头状瘤病毒感染是宫颈癌的主要致病因素,其中受其编码的癌蛋白为 E6,它参与了细胞外信号调节激酶 1/2( extracellular signal-regulated kinase 1/2,ERK1 /2) 信号通路的转导并加剧了正常宫颈上皮的癌变[1].Oishee 等[2]研究发现,E6 蛋白通过 Rap1( Ras-proximate-1 or Ras-related protein 1) 使丝裂原活化蛋白激酶( mitogen-activated protein kinase,MAPK)信号通路激活.同时大量研究表明,基质金属蛋白酶 2( matrix metalloproteinase 2,MMP-2) 、MMP-9 与恶性肿瘤的浸润、转移和发展相关,抑制 MMP-2、MMP-9 的表达是控制癌细胞侵袭和转移及预后的关键[3].黄芩素是中药黄芩的主要成分,具有抗炎、抗氧化、免疫调节以及抑制肿瘤等作用[4],黄芩水提物可以明显抑制宫颈癌 HeLa 细胞的体外生长,与药物的作用时间和浓度成正向关系[5].本研究通过前期实验确定黄芩素处理的时间,用不同浓度的黄芩素处理HeLa 细胞,探索 MMP-2、MMP-9 活性变化和 ERK1 /2表达变化及可能的机制,力图阐明黄芩素处理后MMP-2、MMP-9 活性与 ERK1 /2 信号通路之间的相互关联,为研究宫颈癌的发生、发展提供实验依据.
1 材料和方法
1. 1 材料 HeLa 细胞株由泸州医学院附属医院医学实验中心惠赠,DMEM 高糖培养基购于 HyClone 公司,黄芩素购于Chengdu Must Bio-Techology 公司,其余 PAGE 胶成分购于上海生工公司,明胶粉剂购于 Sigma 公司,小鼠抗 β-actin 抗体及 5 × 上样缓冲液购于碧云天公司,蛋白酶抑制剂购于 Roche公司,兔抗人 MMP-2 抗体、兔抗人 MMP-9 抗体、兔抗人ERK1 /2 抗体及兔抗人磷酸化的 ERK1 /2 ( p-ERK1 /2) 抗体均购于 CST 公司,反转录 PCR 试剂盒购于天根公司.ERK1/2、MMP-2、MMP-9 引物均由上海生工合成.
1. 2 方法
1. 2. 1 细胞培养 用含有 100 mL / L 灭活胎牛血清及 100 U/ mL青霉素、100 μg/mL 链霉素的 DMEM 高糖培养基培养HeLa细胞.置于 37℃、50 mL/L CO2细胞孵箱常规培养.
1. 2. 2 细胞分组 对数生长期细胞,经 2. 5 g / L 胰酶消化后接种( 接种1. 5 ×106个/皿) 到10 cm 直径的一次性培养皿中,使用含 100 mL/L 胎牛血清的 DMEM 高糖培养基培养细胞,等待细胞贴壁,生长 24 h.换无血清 DMEM 高糖培养基同步化处理 24 h 后,用含有( 0、100、200、300) μmol/L 黄芩素的无血清 DMEM 高糖培养基培养 12、24、48 h.镜下观察,与空白对照组相比较,3 个浓度组在 12 h 时,细胞形态和数量变化不大; 24 h 时,300 μmol/L 黄芩素处理组及 48 h 时各浓度处理组,细胞大量凋亡( 凋亡率约 80% ~90%) ,无实验意义,最终选择处理 24 h,空白对照组、100 μmol/L 黄芩素处理组及200 μmol/L 黄芩素处理组.每组设 3 个复孔.分为空白对照组: 不含黄芩素的无血清 DMEM 高糖培养基培养细胞;100 μmol / L 黄芩素处理组的黄芩素终浓度为 100 μmol / L;200 μmol / L黄芩素处理组的黄芩素终浓度为 200 μmol / L.
1. 2. 3 明胶酶谱法检测 MMP-2、MMP-9 的活性变化 黄芩素处理 24 h 后,在相同条件下培养并同步化细胞后,空白对照组加入无血清 DMEM 高糖培养基,100 μmol/L 黄芩素处理组加入含有黄芩素终浓度为 100 μmol/L 的 DMEM 高糖培养基,200 μmol/L 黄芩素处理组加入含有黄芩素终浓度为200 μmol / L的 DMEM 高糖培养基.各组均刺激 24 h 后,收集上清,分装后置于 -80℃保存.每组取 15 μL 上清与 5 μL 上样缓冲液均匀混合,放置于室温 10 min 后,电泳; 电泳结束后把胶放入含 Triton X-100 的复性液中复性40 min,然后轻柔漂洗胶40 min,放入孵育液中,37℃过夜.次日,考马斯亮蓝染色 1 h 后,脱色 30 min.放入成像仪中成像后用 QuantityOne 软件半定量分析图片.通过吸光度( A) 值反应各组金属基质蛋白酶活性的变化.
1. 2. 4 反转录 PCR 法检测 ERK1 /2、MMP-2 和 MMP-9 mRNA的表达 按照反转录 PCR 试剂盒上操作步骤进行 PCR 实验.ERK1 /2 序 列 为 F: 5'-GATTGCCGATCCTGAGCATGAC-3',R: 5'-GCAGGTCAAGGGCCAGAATG-3' ; MMP-9序列为F: 5'-TT-CAGGAGACGCCCATTTC-3',R: 5'-GTCGTCGGTGTCGTAGTT-GG-3'; MMP-2 序列为: F: 5'-AGAGTGCATGAACCAACCAG-3',R: 5'-GCACAAACAGGTTGCAGCTC-3'; β-actin 序 列 为F: 5'-TTTCTTGACAAAACCTAACTTGCG-3',R: 5'-GGACTTC-CTGTAACAATGCATCTC-3',采用独立培养的不同批次细胞重复 3 次实验,取平均值.
1. 2. 5 Western blot 法检测 p-ERK1 /2、MMP-2、MMP-9 的表达 收集好上清的 HeLa 细胞用预冷无菌 PBS 液冲洗 3 次后,加入提前 10 min 混有蛋白酶抑制剂的裂解液提取蛋白,提取后的蛋白经过沸水变性 5 min 后,待温度降至室温,进行100 g / L的SDS-PAGE,电泳结束后进行湿转,转移至 0. 2 μmPVDF 膜上,50 g / L 脱脂奶粉封闭 1 h,加入小鼠抗 β-actin 抗体( 1∶2 000) 、兔抗人 ERK1/2 抗 体 ( 1 ∶ 1 000) 及 兔 抗 人p-ERK1 /2 抗体( 1∶1 000) 、兔抗人 MMP-2 抗体( 1∶1 000) 、兔抗人 MMP-9 抗体( 1∶1 000) ,4℃孵育过夜.次日取出,充分漂洗2 次,每次 20 min 后,加入 HRP 标记山羊抗兔 IgG( H + L) 二抗( 1∶2 000) 或 HRP 标记山羊抗小鼠 IgG( H + L) 二抗( 1∶2 000) ,室温孵育 1 h,再次充分漂洗 4 次,每次 5 min,放入 Bio-Rad 成像仪进行成像并使用 Quantity One 软件分析结果.以目的蛋白与内参 β-actin 的吸光度比值表示为目标蛋白的相对表达量.实验重复 3 次.
1. 2. 6 统计学分析 使用 SPSS16. 0 统计学软件包做统计学分析.计量资料采用 x ± s 表示,采用单因素方差分析,组间两两比较采用 LSD 分析.P <0. 05 表示差异有统计学意义.
2 结果
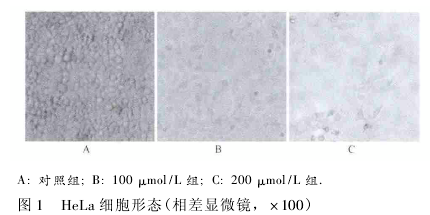
2. 1 细胞形态及数量的变化 经过前期细胞培养筛选出空白对照组、黄芩素处理 24 h 后的 100 μmol/L组和 200 μmol/L 组,镜下观察,随着黄芩素处理浓度从( 0、100、200) μmol/L 逐渐增加,与对照组相比,细胞形态从不规则类圆形变成不规则长梭形,形态由圆形密集排列,变为多角形、细胞稀疏,并且细胞数量明显减少( 图 1) .表明细胞生长受到抑制,且抑制程度与黄芩素的浓度相关.
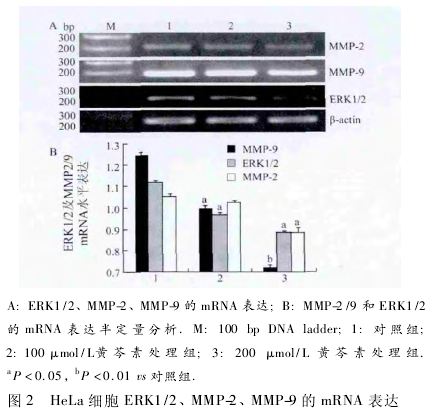
2. 2 黄 芩 素 下 调 HeLa 细 胞 ERK1 /2、MMP-9、MMP-2 mRNA 的表达 在黄芩素处理 24 h 后,ERK1 /2、MMP-9、MMP-2 的 mRNA 水平随黄芩素的浓度增加逐渐降低,与对照组相比,100 μmol/L 黄芩素处理组表达均减弱( ( P <0. 05,图 2) ; 200 μmol/L黄芩素处理组表达明显下降,ERK1/2、MMP-2、MMP-9的 mRNA 水平明显降低,差异有明显统计学意义( P <0.01,图2) .
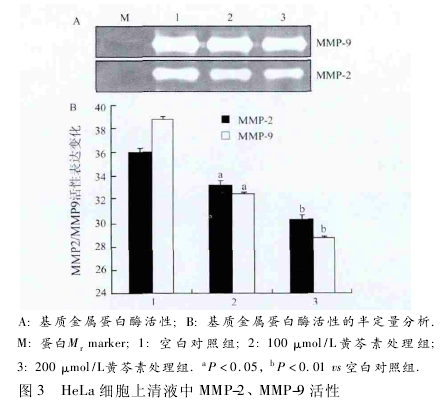
2. 3 黄芩素降低 HeLa 细胞 MMP-2、MMP-9 活性在黄芩素处理24 h 后,MMP-9 和 MMP-2 活性与表达随黄芩素的浓度增加逐渐降低,与对照组相比较,100 μmol / L 黄芩素处理组表达减弱 ( P < 0. 05,图 3) ; 200 μmol/L 黄芩素处理组表达明显减弱,MMP-2 和 MMP-9 活性明显受到抑制,差异有统计学意义( P <0. 01,图 3) .
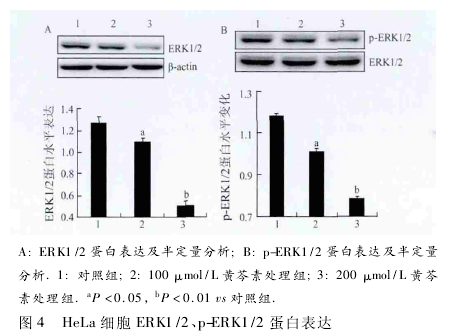
2. 4 黄 芩素下调 HeLa 细胞 MAPK 信号通路中ERK1 /2、p-ERK1 /2 蛋白表达 在黄芩素处理24 h后,ERK1/2、p-ERK1/2 随黄芩素的浓度增加而逐渐降低,与对照组相比,100 μmol/L 黄芩素处理组表达下降( P <0. 05,图 4A、B) ,200 μmol/L 黄芩素处理组下降峰明显,差异有统计学意义( P < 0. 01,图4A、B) .
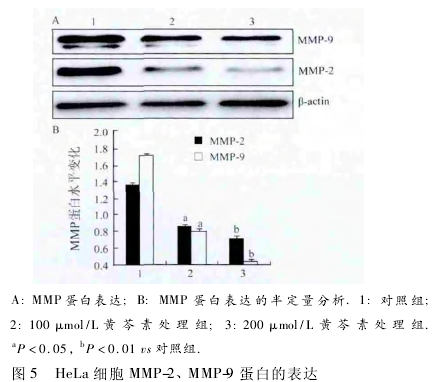
2. 5 黄芩素下调 HeLa 细胞 MMP-9 和 MMP-2 蛋白的水平 采用 Western blot 法检测 MMP-9 和 MMP-2蛋白的表达.在黄芩素处理 24 h 后,MMP-9 和MMP-2 随黄芩素的浓度增加逐渐降低,与对照组相比,100 μmol/L 黄芩素处理组表达下降( P < 0. 05,图 5) ,200 μmol/L 黄芩素处理组明显下降,差异有统计学意义( P <0. 01,图 5) .
3 讨论
宫颈癌的发生、发展是由 HPV 感染后引发内源性因素而引起的,相关癌基因被激活、抑癌基因失活以及机体免疫调节机制失衡等引起细胞增殖与凋亡的异常,导致组织癌变[6 -7].黄芩茎叶总黄酮对HeLa细胞增殖具有显着的抑制作用[8],ERK1/2 和p-ERK1 /2 在宫颈癌的高表达在其恶变、侵袭过程中发挥了重要的作用,并可能是宫颈癌患者预后不良的指标之一[9].ERK1/2 是 MAPK 的重要组成部分,通过磷酸化激活从而有效控制细胞增殖等[10],本实验结果证实在宫颈癌中黄芩素能降低 ERK1/2 和p-ERK1 /2的表达,抑制 HeLa 细胞的增殖,与其他研究结论一致.Wu 等[11]研究表明黄芩素可抑制皮肤癌肿瘤细胞的侵袭和转移.若能有效抑制与肿瘤侵袭相关的因子 MMP-2、MMP-9 的表达,势必会延缓肿瘤的侵袭与转移.Wang 等[12]研究表明,黄芩素可以有效抑制 MDA-MB-231 细胞的黏附、转移及侵袭,这种作用可能是通过抑制 MMP-2、MMP-9 的分泌与表达,同时可能通过抑制 MAPK 信号通路实现的.
黄芩素对宫颈癌 HeLa 细胞的抑制作用是否涉及MMP-2、MMP-9 的改变及 MAPK 信号通路的激活?
本实验不仅证明在宫颈癌中黄芩素抑制了 MMP-2、MMP-9 的表达,同时也证实,在此过程中 MAPK 信号通路被激活并受到抑制.研究发现,多种肿瘤中MMP 的表达明显高于正常组织,且与肿瘤的侵袭和转移相关[13 -15],是否可以通过抑制 MMP 家族中与侵袭相关的关键因子 MMP-2、MMP-9 活性达到抑制宫颈癌 HeLa 细胞的增长? 本实验证实黄芩素能够通过抑制 MMP-2、MMP-9 的表达,达到抑制肿瘤的作用.
本研究结果表明: 不同浓度的黄芩素处理组MMP-2、MMP-9 及 p-ERK1 /2 较对照组表达降低.黄芩素作用于宫颈癌 HeLa 细胞导致 ERK1/2 表达降低,说明 ERK1/2 信号通路参与了相关的作用并被激活.
p-ERK1 /2 表达的降低可能与黄芩素下调 ERK1 /2 有关.MMP-2、MMP-9 的活性、mRNA 及蛋白水平的表达随黄芩素浓度增加而逐渐降低表明,黄芩素可能通过抑制 MMP-2、MMP-9 活性来改变其表达,从而抑制宫颈癌 HeLa 细胞的增殖,最终有效抑制宫颈癌的发展.
综上所述,黄芩素能够通过激活 MAPK 信号通路中 p-ERK1/2,降低 MMP-2 和 MMP-9 的活性,阻止宫 颈 癌 HeLa 细 胞 的 生 长.提 示 阻 滞 或 拮 抗ERK1 /2 及抑制 MMP-2、MMP-9 活性,有助于阻止、延缓宫颈癌的发生和发展,改善其预后,为临床宫颈癌治疗提供新的思路.
参考文献:
[1]李 刚,姚珍薇. ERK1/2 信号转导通路在宫颈癌发病中的作用[J]. 山东医药,2007,47( 24) : 118 -119.
[2]Chakrabarti O,Veeraraghavalu K,Tergaonkar V,et al. Human papilloma-virus type 16 E6 amino acid 83 variants enhance E6-mediated MAPKsignaling and differentially regulate tumorigenesis by Notch signalingand oncogenic Ras[J]. J Virol,2004,78( 11) : 5934 - 5945.
[3]Song C,Zhu S,Wu C,et al. Histone deacetylase ( HDAC) 10 suppressescervical cancer metastasis through inhibition of matrix metalloproteinase( MMP) 2 and 9 expression[J]. J Biol Chem,2013,288( 39) :28021 - 28033.
[4]许杜娟,陈敏珠. 黄芪总提物抗肿瘤作用及其机制研究[J]. 中华中医药杂志,2006,21( 12) : 771 -772.
[5]王晓莉,苏志红,李妍芹,等. 黄芪对体外培养人宫颈癌 HeLa细胞的抑制作用[J]. 西北国防医学杂志,2010,31( 4) : 270 -272.
[6]张晓金,归绥琪. 宫颈癌发病机制的研究进展[J]. 中国妇幼健康研究,2008,19( 1) : 56 -59.
[7]比丽克孜. 艾克木,封 敏,李秀梅,等. MTA1、MMP2、MMP7在宫颈癌中的表达及其意义[J]. 中国现代医学杂志,2013,23( 29) : 51 - 54.
[8]郭慧芳,徐 倩. 黄芩茎叶总黄酮对 HeLa 细胞增值的影响[J].承德医学院学报,2013,30( 4) : 275 -277.
[9]章丽霞,刘 钧,黄一凡,等. ERK1/2 和 P-ERK1/2 蛋白在宫颈癌中的表达和意义[J]. 现代肿瘤医学,2012,20 ( 2) :353 - 356.
[10]任婷婷,贺红焰,喻小兰,等. 血管紧张素-( 1-7) 抑制低氧诱导的大鼠肾小管上皮细胞转分化[J]. 细胞与分子免疫学杂志,2013,29( 6) : 593 - 596.
[11]Wu B,LI J,Huang D,et al. Baicalein mediates inhibition of migrationand invasiveness of carcinoma through Ezrin in A431 cells[J / OL].BMC Cancer,2011,11: 527.
[12]宋 波,金锡御,潘进洪,等. MMP-2 及 MMP-9 在膀胱癌中的表达及在侵袭转移中的作用[J]. 中华泌尿外科杂志,1999,20( 7) : 15 - 17.
[13]Zhang M,Zhu GY,Gao HY,et al. Expression of tissue level of matrixmetalloproteinases and tissue inhibitors of metalloproteinases in gastricadenocarcinoma[J]. J Surg Oncol,2011,103( 3) : 243 - 247.
[14] Wang L,Ling Y,Chen Y,et al. Flavonoid baicalein suppressesadhesion,migration and invasion of MDA-MB-231 human breastcancer cells[J]. Cancer Letts,2010,297( 1) : 42 - 48.
[15]祝立和,卢洪胜,周凯敏,等. STAT3 基因与 MMP-2、MMP-9 在乳腺浸润性导管癌中的表达及其相互关系[J]. 中国现代医生,2012,50( 29) : 58 - 61,64.





