
树木在生长过程中,树轮特征能够真实地记录并反映周围环境特征。在众多环境因素中,水分是影响树木树轮生长的关键因子之一。提取记录在年轮内部的更详细的环境信息,有助于进一步明确树轮特征和环境之间的关系。古尔班通古特沙漠是我国最大的固定与半固定沙漠。受沙土物质组成和沙垄部位对水分再分配的影响,不同沙垄部位土壤水分和植被生长特征有较大差异。
白梭梭( Haloxylon persicum) 为梭梭属( Haloxylon) 藜科( Chenopodiaceae) 植物,灌木或小半乔木,是亚洲荒漠区分布最广的荒漠植被类型。白梭梭具有防风固沙、耐旱、耐盐碱等特点,是我国干旱荒漠地区的主要固沙造林的优良先锋树种和生物量较高的沙漠植物群落重要建群种之一,对维护脆弱荒漠生态系统的稳定性具有重要意义。由于其材质坚硬,切片困难且易碎,不易得到完整的较大的切片图像,解剖结构不清楚,有关白梭梭树轮解剖特征鲜见报道。文中对古尔班通古特沙漠三种沙垄部位白梭梭组织比量进行了研究,初步分析了树轮组织比量特征,为白梭梭种群保护和恢复等研究提供理论依据。
1 材料与方法
1. 1 采样区概况
材料采集地在古尔班通古特沙漠腹地农八师 148 团石西公路南侧,地理位置 N44°00. 269'E86°17.383',海拔 383m,属于温带荒漠气候,夏季炎热,冬季寒冷。平均气温 4 ~ 6℃ ,年降水量仅 115mm,蒸发量1942mm。荒漠植被梭梭( Haloxylon ammodendron) 和白梭梭为该区的建群种。沙漠内部植被覆盖率 15%~ 30% 。固定沙丘和半固定沙丘占整个沙漠面积的 96% 。
1. 2 采样方法
2012 年 10 月 28 日于农八师 148 团石西公路南侧沙漠中选取迎风坡、坡顶、背风坡三种沙垄部位长势良好的白梭梭株丛,选取外观良好,无明显人为损伤,胸径基本一致的白梭梭树干( 一级枝) ,自过渡茎锯掉,带回实验室,在树干的基部、中部、顶部锯下圆盘,部分圆盘置于 FAA 固定液中保存,部分圆盘待打磨使用。
1. 3 样品处理
将待打磨的圆盘按照锯面和树干轴向垂直锯好,用不同粒度规格的砂纸做预处理打磨,之后用粒度为1500#规格的砂纸做最后处理。
把用 FAA 固定液保存的圆盘锯成长度约 5cm,沿髓心用斧头劈开,从圆盘髓心到边缘沿半径方向均匀选取三个点,把圆盘劈成小木条并标记,断面大小以易于切片为宜,同一个圆盘的相同半径位点可以多劈几个木条,以备软化切片使用。迎风坡、坡顶、背风坡三种沙垄部位,每种沙垄部位锯圆盘三个,分别为树干基部圆盘、中部圆盘、顶部圆盘,每个圆盘从髓心到边缘,可劈为髓心、中部和边缘三种小木条,因此三种沙垄部位共有不同木条组 27 组。把 27 组小木条分别编号,放置于小烧杯中,加入适量酒精 - 甘油混合液,浸泡、加热以软化木条,达到切片要求即可,软化好的木条可以放置酒精 - 甘油混合液中长期保存。
1. 4 测定方法
1. 4. 1 年轮特征测定
打磨好的圆盘置于 EPSON Scan V700 扫描仪,选择 6400dpi 进行扫描,得到的图像用 WinDENDRO 树轮分析系统处理,通过木射线相交确定髓心,每张图像测定 4 组数据,得到不同沙垄部位白梭梭的树轮宽度和深棕色带宽度值,尔后求均值以分析。
1. 4. 2 组织比量测定
取出软化好的小木条,擦拭表面的混合液,用锋利的切片刀进行切片,切片放置到铺有少量甘油的载玻片上,用解剖针展平,置于显微镜下观察,并用显微镜配套的微距相机拍照。选取点计数法测定组织比量,每试样测定不少于 30 次。
2 结果与分析
2. 1 白梭梭树轮特征
打磨好的圆盘可以看到明显的木射线。白梭梭树轮生长带由深棕色带和浅棕色带组成,深棕色带与浅棕色带形成带状相间分布( 图 1) 。浅棕色带部分深入深棕色带中,形成楔形集合体,与轴向薄壁组织为主组成的浅棕色带联接形成一条完整生长带。白梭梭连续形成层一年多次分裂,连续产生多个同心圆式排列的形成层环,形成次生维管组织,即白梭梭一年形成多个生长带( 树轮) 。该株白梭梭位于沙丘背风坡,最长半径含生长带 36 个,依据宋于洋等的年龄估算模型,可知树龄为 8 年。【图1略】
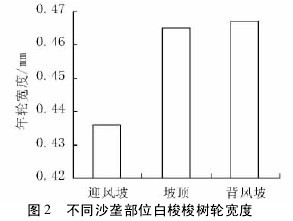
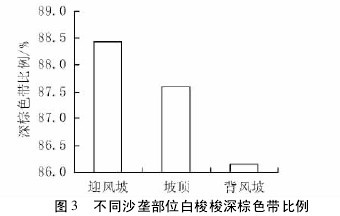
三种沙垄部位下,白梭梭树轮宽度表现出明显的差异性,背风坡和坡顶树轮宽度显著高于迎风坡( 图2,p < 0. 01) 。不同沙垄部位下,白梭梭树轮深棕色带的比例也有较大的差异,迎风坡比例最大,坡顶次之,背风坡最小( 图 3,p <0. 01) 。结合图 2 可以看出,迎风坡白梭梭树轮窄且深棕色带比例较高,而晚材( 白梭梭深棕色部分) 率的大小可以作为衡量木材强度大小的标志,晚材率大的树种,其木材强度也相应地较高,因此迎风坡白梭梭树干坚硬,更有利于抵抗风沙危害。【图2-3】
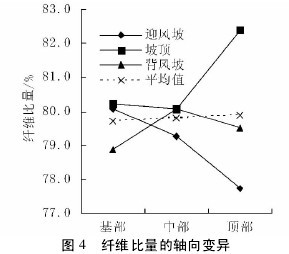
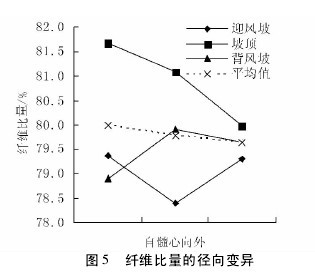
2. 2 不同沙垄部位对纤维比量的影响
图 4 与图 5 表明,不同沙垄部位下的白梭梭纤维比量在轴向和径向上,变异趋势各不相同。迎风坡白梭梭的纤维比量在轴向上呈现下降趋势,而在径向上却是先下降后上升趋势。坡顶白梭梭的纤维比量在轴向上呈现微弱下降而后上升的趋势,在径向上是下降趋势。背风坡白梭梭的纤维比量在轴向和径向上均呈现先上升后下降的趋势。【图4-5】
2. 3 不同沙垄部位对导管比量的影响
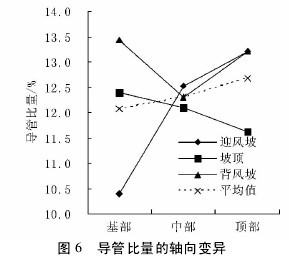
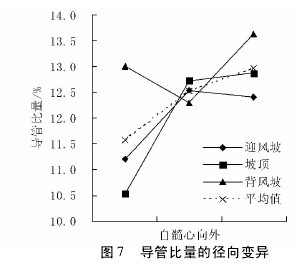
不同沙垄部位白梭梭的导管比量在轴向上,迎风坡呈现上升趋势,坡顶为下降趋势,背风波是先下降后上升,坡顶和背风坡的表现为树干顶部导管比量低于基部,而迎风坡表现为顶部高于基部( 图 6) 。导管比量在径向上,迎风坡和坡顶均表现为上升趋势,背风坡表现为先下降后上升趋势,尽管表现不一致,但是,导管比量边缘部位高于髓心部位,在三种沙垄部位下却表现为一致,说明导管在木材边缘部分含量较高( 图 7) 。【图6-7】
2. 4 不同沙垄部位对木射线比量的影响
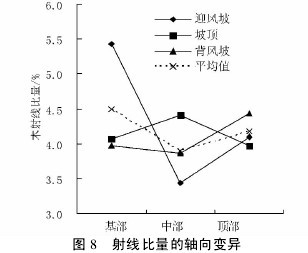
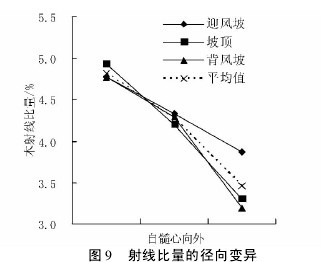
不同沙垄部位白梭梭的木射线比量在轴向的变异趋势各不相同。迎风坡表现为先下降后上升的趋势,而坡顶和背风坡变化幅度不大( 图 8) 。在径向上,三种沙垄部位下的白梭梭木射线比量变异趋势表现一致,均为明显下降趋势( 图 9) ,说明髓心部位木射线含量较高。【图8-9】
2. 5 不同沙垄部位对轴向薄壁组织比量的影响
图 10 与图 11 表明,不同沙垄部位白梭梭的轴向组织比量在轴向的变异趋势为,迎风坡表现为上升趋势,坡顶和背风坡表现为下降趋势。在径向上,迎风坡和背风坡变化幅度很小,而坡顶表现为先下降后上升的趋势。无论是轴向还是径向,轴向薄壁组织比量值,迎风坡高于背风坡,背风坡高于坡顶。【图10-11】
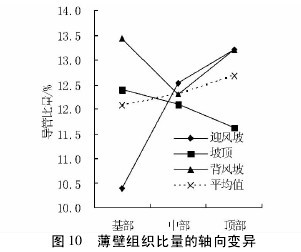
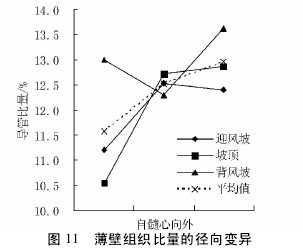
2. 6 不同沙垄部位对组织比量的影响
白梭梭解剖结构中,深棕色生长带主要由纤维、射线和导管构成。浅棕色带主要由轴向薄壁组织和导管构成( 图 12) 。坡顶白梭梭的纤维比量最高,迎风坡和背风坡相差不大( p >0. 01) 。迎风坡和坡顶白梭梭的导管比量没有显著差异( p >0. 01) ,但都小于背风坡的导管比量( p <0. 01) 。三种沙垄部位下,白梭梭的木射线比量和轴向薄壁组织没有显著差异( p >0. 01) 。但表现出迎风坡木射线比量和轴向薄壁组织比量最高趋势,分别为 4. 32%和 4. 61%( 图 13) 。【图12略.图13】
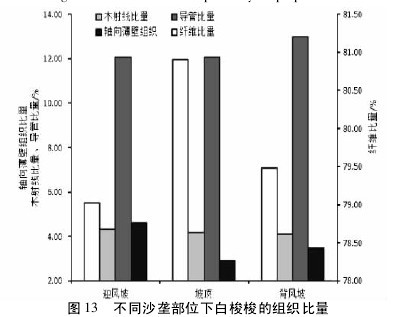
3 讨论
本研究中,迎风坡白梭梭树轮密而且深棕色带比例较大,材质密实坚硬,坡顶次之,背风坡最差。试验中发现,对于半固定沙丘,背风坡白梭梭存在沙埋现象,坡顶和迎风坡白梭梭常因为缺水或者其他原因生长状态差甚至干枯死亡,迎风坡白梭梭枝干树轮常偏心生长,呈现单侧生长的现象,据王炜、梁存柱等的研究,推断是由荒漠区风沙流和强烈的地表辐射所致。某些迎风坡梭梭的周皮不发达,在生长受限制一侧已开始损失木质部,有的髓心坏死甚至中空,推测这也是部分白梭梭树干甚至株丛枯死、折断的原因之一。
白梭梭纤维比量在不同沙垄部位下没有明显的变异规律,在轴向上呈现上升趋势,径向上呈现下降趋势。这与费本华对铜钱树的研究结果恰恰相反,研究表明,铜钱树纤维比量随轮龄的增大初始为递增,约在 14 年后逐渐变缓; 在轴向上,纤维比量自基部向上逐渐减小。胡进波等( 2008) 对尾巨桉的研究显示,纤维比量在从髓心向外的数个生长轮内变异规律不明显,部分家系在近树皮处纤维比量呈减小趋势,原因可能是由于幼年期形成层原始细胞快速分裂形成的导管细胞较少,形成层后增长作用较弱,促使纤维细胞比量相对多一些; 进入成熟期后,形成层原始细胞分裂趋于稳定,管孔的形成层后增长作用加强,促使纤维细胞比量相对的要少一些。苏文会等( 2005) 对大木竹的研究显示,纤维比量随秆的高度增加而增大,这与文中研究结果一致。
不同沙垄部位平均导管比量,在轴向和径向上均呈现上升趋势。费本华对铜钱树的研究显示,轴向上导管比量在逐渐增加,径向上导管比量缓慢递减。卢翠香等( 2012) 的研究显示,马尾松管胞比量随树龄增加呈波动减小,而随树干高度的增加,呈现增加 - 减小 - 增加规律。两者在导管比量径向变化的结果一致,与文中研究结果相反,推测原因可能是木质部母细胞分裂必需的生长因子差异分布的结果,在幼年期,细胞分裂快,导管分子所需的生长因子受到抑制,导管分子的分化较弱,比量较小; 到成熟期后,细胞分裂趋于稳定,良好的细胞迅速扩展,并分化为导管分子,故导管比量相对较多。
不同沙垄部位平均木射线比量,在轴向上是先减小后增加的趋势,而径向上,明显表现为下降趋势。
木射线可从任一树轮处发生,一旦发生,它随着直径的增大而延长,直到形成层。费本华( 1994) 的研究表明,铜钱树在径向,随轮龄的增大木射线比量初始为递减,后逐渐变缓; 在纵向,木射线比量自基部向上逐渐减小。铜钱树木射线比量在径向上的变化趋势与白梭梭基本一致。马顺兴等( 2006) 的研究表明,日本落叶松无性系木材木射线比量由髓心向外,开始迅速下降,到第 2 年轮后缓慢波动下降或上升。究其原因,前人研究认为木射线组织比量的大小与纤维比量和导管比量大小有很大关系,随着年龄的增加,导管细胞形成层增长加强,导管比量增大,致使木射线和纤维比量相应都有所减小。
不同沙垄部位平均轴向薄壁组织比量,轴向变化趋势是先上升后下降,径向变化趋势是先下降后上升。汪佑宏等( 2000) 的研究表明不同淹水程度对长江滩地枫杨轴向薄壁组织比量的影响,由髓心向外是先减少后又回升,呈"V"形,自基部向上先是急剧增加后又急剧减少,最后又呈波动性地增加。这与文中研究的变化趋势相一致。对巨桉家系木材研究表明,轴向薄壁组织比量径向上的变异无一定的规律,波动性较大,并且尾巨桉各家系轴向薄壁组织的变异性大。
统计得出,白梭梭纤维比量为 79. 80%,导管比量为 12. 36%,木射线比量为 4. 19%,轴向薄壁组织比量为 3. 66%。对生长在热带海洋潮间带的红树植物的研究表明,木本植物白骨壤的木纤维比量、木射线比量、导管比量、薄壁组织比量平均值分别为 71%、7%、22%、1%。对欧美杨 107 杨木材解剖表明,其木射线比量变化幅度为 10. 21 ~13. 60%,导管比量为 17. 30 ~24. 40%,纤维比量为 64. 37 ~71. 24%。
对铜钱树的研究表明,其组织比量为: 纤维比量69. 4%,导管比量11. 4%,木射线比量16. 9%,薄壁组织比量 2. 3%。对比以上几种植物,白梭梭的纤维比量最高,这或许可以解释为什么白梭梭的材质异常坚硬。另外,白梭梭发达的机械组织可减少同化枝萎蔫时的损伤,增强对荒漠生境干旱胁迫的抵御能力。
不同沙垄部位中,坡顶白梭梭的纤维比量最高( 80. 90%) 。这可能是在沙垄各个部位中,坡顶风速最大,生长在坡顶的白梭梭长期适应风吹产生应激性反应,导致其担负机械支持作用的纤维比量增大。
生长在不同生境中植物表现出结构上的差异,通常被认为是对特定生境的进化适应。植物导管通过调节直径大小来适应水分变化。由于地形对土壤水分的再分配及土壤蒸发差异,三种沙垄部位中,迎风坡和坡顶土壤含水量低于背风坡,白梭梭采取减少导管比量来适应较低的土壤水分条件,因而背风坡导管比量最高。三种沙垄部位下,白梭梭的木射线比量和轴向薄壁组织比量没有显著差异( p > 0. 01) 。
这与符韵林( 2003) 对杉木木材解剖的研究结果一致。
4 结论
( 1) 三种沙垄部位白梭梭的树轮宽度,背风坡高于坡顶高于迎风坡; 深棕色带比例,迎风坡高于坡顶高于背风坡。( 2) 白梭梭纤维比量、导管比量、木射线比量、轴向薄壁组织比量分别为 79. 80%、12. 36%、4. 19%、3.66% 。三种沙垄部位中,坡顶白梭梭的纤维比量最高,背风坡导管比量最高,木射线比量和轴向薄壁组织比量在各沙垄部位间没有显著差异。( 3) 纤维比量从树干基部到顶部缓慢增大,自髓心向外呈减小趋势,变幅不大。导管比量从树干到基部、自髓心向外均呈现增大趋势,变幅不大。木射线比量从树干基部到顶部,随高度的增加先减小,达到一定值后增大,自髓心向外呈现减小趋势。轴向薄壁组织从树干基部到顶部,随高度的增加先增大后减小;自髓心向外,呈现先减小后增大的趋势。





