
喜旱莲子草 Alternanthera philoxeroides 属于苋科 Amaranthaceae 莲子草属 Alternanthera, 原产南美洲, 主要生长于各种淡水生态系统的水陆交界区域和农田等, 由于其茎叶繁茂, 根系发达, 生长速度迅猛, 繁殖能力惊人, 具有很强的适应环境及入侵特性, 后被传播到北美洲、 大洋洲、 亚洲等地, 逐渐成为重要入侵植物, 也是 中国国家环境保护总局公布的 9 种危 害最大的外来入侵植物之一 , 已经对入侵地生态系统及生物多样性构成了严重威胁。 该入侵种能够抑制和排挤乡土植物, 阻塞航运, 污染环境, 引发一系列不良的生态学后果。 近年来, 该入侵种在东苕溪江被陆续发现, 其入侵面积和个体数量不断增长。 苕溪江是浙江八大水系之一, 是中国东南沿海和太湖流域唯一一条没有独立出海口的南北向的天然河流。 作为太湖流域的主要水源和杭州饮用水源之一, 东苕溪江发源于临安市临目乡马尖岗,向东流经临安市的里畈、 桥东、 临天、 青山和余杭镇, 自余杭镇折北而流。 喜旱莲子草在东苕溪江的继续扩散, 将严重威胁苕溪流域的供水安全和水生态安全。 基于此, 本研究对东苕溪江不同流域段喜旱莲子草群落特征进行了调查, 分析喜旱莲子草对入侵地植物多样性的影响, 为入侵地物种多样性的保护提供参考。
1、 材料与方法
1.1 样地选择
本研究于 2012 年 7-9 月在东苕溪江不同河段进行。 从源头开始, 至末段杭州余杭安溪镇段。 调查地点选择受不同程度干扰的河流段, 分别选取河流上游源头、 中游和下游附近进行采样, 共选取临安市太湖源(指南村)、 临目乡(里畈水库上游)、 里畈水库下游、 东湖村, 及杭州市余杭区余杭镇、 陡家门、 瓶窑镇和安溪镇 8 个调查点。 调查地点见图 1。

1.2 研究方法
采用群落调查法, 对不同调查地点喜旱莲子草群落物种多样性进行调查, 分析群落内植物的种类、 数量和盖度等。 具体操作为: 在调查地点随机选定样地中心点, 设置 10.0 m × 10.0 m 正方形样区, 并在其内设置 5 个 0.5 m × 0.5 m 的样方, 取样点采取 “W” 型排列。 调查记录各个样方中喜旱莲子草及其他植物种类、 数量、 盖度, 并记录样方内喜旱莲子草的分枝数、 茎长、 叶面积和节间距。 其中节间距为茎长与枝条节数的比值。 喜旱莲子草的株数(丛数)以地下根状茎来区分, 枝条在同一根状茎则视为 1 株。
喜旱莲子草叶面积的计算参照张震等的方法, 计算公式为: s=π×(a/2)×(b/2)。 其中, a 为叶宽(cm), b 为叶长(cm), 长和宽均为每分枝第 2 节(近地端)上叶片的长轴和最宽处测量所得。 重要值的计算公式为: 重要值=(相对密度+相对频度+相对盖度)/3。 其中: 相对密度=(样方内某种植物的株数/样方内所有植物的株数之和)×100%; 相对频度=(每个样点中出现某植物的样方数/该样点中总的样方数)×100%; 相对盖度=(样方中某种植物的分盖度/样方中所有植物的分盖度之和)×100%。 群落生物多样性指数的计算参照下列公式: Simpson 多样性指数

Pielou 均匀度指数(JH): JH=H / lnS。 其中, Pi为第 i 个物种个体数占群落中总个体数的比例, S 为物种总数。
1.3 数据分析
采用 SPSS 统计软件对调查数据进行单因素方差分析, 并对不同采样点指标进行差异显著性检验。
2、 结果与分析
2.1 不同河流段生境喜旱莲子草形态特征
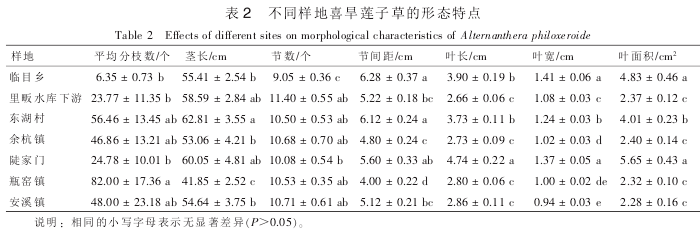
受人类干扰程度不同, 不同调查点喜旱莲子草呈现不同分布特征。 在太湖源源头地区未发现喜旱莲子草的分布, 其他地点均发现该入侵种的分布(表 1)。 不同采样点生境受干扰程度不同, 喜旱莲子草形态差异也较大(表 2)。 其中, 瓶窑镇样地内喜旱莲子草植株的分枝数最多, 平均为 82 个·株-1, 临目乡样地喜旱莲子草植株分枝数最小, 仅 6.35 个·株-1。 东湖村、 陡家门和里昄水库下游采样点的喜旱莲子草植株的茎长最大, 并且后 2 个采样点喜旱莲子草茎长与其他采样点相比差异显著(P<0.05)。 各采样点平均节数较多, 瓶窑镇采样点节间距最短, 为 4.00 cm。 不同采样点叶长差距比较明显, 最长为 4.74cm, 最短为 2.66 cm。 里畈水库下游、 余杭、 瓶窑镇和安溪镇采样点喜旱莲子草植株的平均叶面积较小,为 2.28~2.40 cm2; 而其他采样点该入侵种的平均叶面积较大, 为 4.01~5.65 cm2。

2.2 不同样地喜旱莲子草的分布及对植物多样性的影响
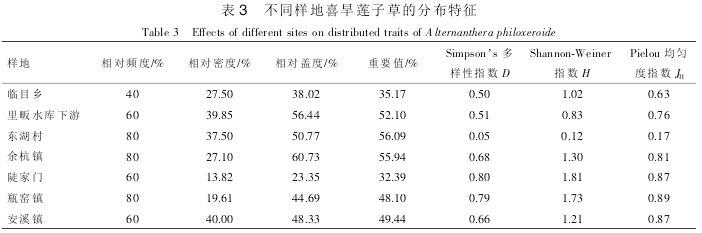
喜旱莲子草在东湖村段、 余杭镇以及瓶窑段出现的相对频度最高, 均为 80%。 安溪镇和里畈水库下游该植物相对密度最大, 分别为 40.00%和 39.85%。 余杭镇采样点喜旱莲子草相对盖度最大, 为60.73%; 除去未发现该入侵种的太湖源样点, 喜旱莲子草相对频度和相对盖度最小值均出现在陡家门采样点(表 3)。 另外, 据相关分析, 喜旱莲子草茎长、 节间距与重要值显著相关(相关系数分别为 0.832 和0.765, P<0.05)。
不同采样点喜旱莲子草的密度大小对其伴生物种影响较大(表 4)。 安溪镇、 里畈水库下游和东湖村段采样点(相对密度分别为 40.00%, 39.85%和 37.50%), 喜旱莲子草相对密度较大, 重要值较高, 其伴生植物种类较少, 分别为 4 种、 3 种和 2 种; 陡家门采样点喜旱莲子草密度较小(13.82%), 重要值较低, 伴生植物种类最多(8 种)。 东湖村采样点喜旱莲子草重要值最高, 伴生物种数最少(2 种), 且大部分伴生植物在生长期却呈现逐渐枯萎趋势, 喜旱莲子草为该样地中的优势物种。 余杭镇采样点喜旱莲子草重要值也较高, 伴生植物为 5 种。 喜旱莲子草伴生植物的种数和数量与喜旱莲子草的数量和重要值之间并无明显的相关性(P>0.05)。
随着喜旱莲子草重要值的升高, 群落 Simpson 多样性指数呈下降趋势, 如东湖村, 多样性指数仅为0.05, 而陡家门采样点 Simpson 多样性指数为 0.80, 表明随着喜旱莲子草种群扩散, 其他物种逐渐受到排挤。 Shannon-Weiner 指数也呈现相同的变化趋势。 伴生植物种类与 Simpson 多样性指数、 Shannon-Weiner 指数和 Pielou 均匀度指数显著相关(相关系数分别为 0.902, 0.958 和 0.829, P<0.05), 在伴生植物种类较少的取样点, Pielou 均匀度指数较低, 说明喜旱莲子草的扩散降低了当地的物种多样性。
3、 结论与讨论
研究表明: 喜旱莲子草的分布特征与东苕溪不同河流段人类活动干扰密切相关。 在东苕溪上游人类干扰弱的地区, 喜旱莲子草的分布较少, 其相对盖度和重要值都较低。 在中游人类活动密集的河段, 喜旱莲子草分布较多。喜旱莲子草在不同生境中可表现出不同的形态特征, 如在水分适宜的生境中茎叶营养生长旺盛。
本研究结果也表明: 在苕溪江沿岸, 如陡家门和临目乡, 各采样点周围水分适宜, 喜旱莲子草茎较长、节间距较大、 叶面积较大、 分枝数相对较少。 这是因为湿生环境中的喜旱莲子草较长的茎能够增强植株的水分疏导能力, 提高占据养分空间的能力。 据相关研究: 喜旱莲子草入侵后, 能通过快速分枝使盖度不断增加, 排挤本地物种, 导致群落物种丰富度明显降低, 在水库边、 河道内形成优势种群, 并致使此类生境中该植物的伴生植物种类和数量减少。 同时, 喜旱莲子草能够产生化感物质抑制周围其他生物生长。 喜旱莲子草密度较大的生境中 , 伴生植物的密度较小 。 当喜旱莲子草的重要值大于 1.5时, 物种丰富度随重要值增加而减小, 喜旱莲子草的入侵对生物多样性产生不利影响。 根据本研究发现, 各采样点喜旱莲子草的重要值均大于 1.5, 表明喜旱莲子草对东苕溪江本地种已产生一定的抑制作用, 降低了生境内植物群落的多样性(东湖村喜旱莲子草重要值最高, 为 56.09%, 其伴生种只有 2 种)。


本研究结果表明: 喜旱莲子草伴生物种的种数和数量与喜旱莲子草的数量和重要值之间并无明显的相关性(P>0.05), 推测其原因可能是由于各个河流段喜旱莲子草的入侵阶段不同。 因此在某些喜旱莲子草重要值高的样地, 虽然喜旱莲子草处在旺盛的生长期, 盖度和重要值都很高, 但是它处在入侵初期阶段, 还未对本地物种造成明显的排挤。 但随着喜旱莲子草的迅猛扩散, 将会对本地物种产生严重威胁, 甚至危及水生态安全。 如果不采取相关措施, 入侵种喜旱莲子草在苕溪江流域有继续扩散的趋势,并将逐渐排挤乡土植物物种, 造成严重的生态学后果, 危及生态系统的平衡。 因此, 必须密切监控该流域喜旱莲子草的扩散, 避免产生更严重的生物安全问题。
参考文献:
[1] 潘晓云, 耿宇鹏, ALEJANDRO S, 等. 入侵 植物喜旱莲子草 ———生物 学 、 生态学及管理[J]. 植物分类学报,2007, 45(6): 884 - 900.





