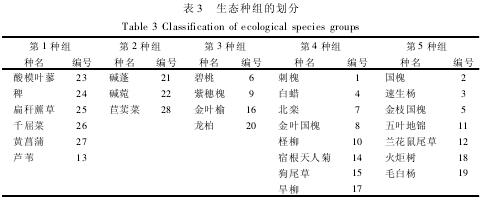
为了更真实的反映群落的种间关联,以Spearman秩相关分析结果为主,参考χ2检验及联结系数、DCA排序距离( 表2) ,将临港湿地植物群落的28个主要物种划分为5个生态种组。种间联结测定值在一定程度上反映了物种间的相互关系以及它们与环境因子的关系,所以可将它们作为生态种组划分的重要参考[6],如表3,第1生态种组主要为湿生植物种组,该组内物种均生活在水分充足地区,多为多年生挺水草本植物,位于积水湿地或潮湿湿润环境处; 第2生态种组为滨水盐生植物种组,三种植物生态位相近,生长环境相似,多为潜流湿地或滨水盐土区域; 第3种组主要为群落演替伴生种,分布广泛,对水分要求不高,临水边坡、小路均有分布; 第4种组主要为湿地绿地组团过渡带物种,常出现在路边,与龙柏、紫穗槐等组成较好的乔灌景观; 与第5种组为受人工干扰较大的旱生植物种,数量较多,分布于湿地周边人工林。
4讨论
结合χ2检验和种间联结系数(OI) ,在χ2中13对呈现负显着相关的种对,Spearman秩相关系数并不显着,在χ2检验中,显着正相关与极显着正相关的种对在秩相关分析中仍然呈显着与极显着相关。一般认为,χ2检验只是对种间关联性给出一个概括性的结论,依据的2 × 2联列表的数据是定性数据,用1和0表示种在样方中存在于否。在数量数据转化为二元数据过程中就必然会损失一定的信息量,而Spearman秩相关系数依据的是定量的数据,减少了信息量的损失,秩相关分析的灵敏度要远高于χ2检验[18].χ2检验与Spearman秩相关检验中,负关联种对所占比例较大,说明主要物种间对生境具有不同的生态适应性和相互分离的生态位; 原生植物群落多为湿生草本,滨水挺水植物,与人为干扰群落本身生态学特性上存在差异,加之恢复和重建群落演替过程复杂,影响因素多,在长期的发展演替中,物种之间发生分化,分别占据了一定的生态位。因此,负关联种对多,但关联性小,群落稳定性不强。
χ2检验和Spearma秩相关系数都表明,呈显着相关的种对较少,种对间相关性较弱,可能与湿地的生境类型异质性有关,如选取样地环境差异较大,包括河道湿地、表潜流湿地、季节性积水的漫滩、疏林草地、堤岸绿地、河岸及路旁等; 不同地段对应不同的优势种,体现了植物对水分条件的不同响应,如第1种组的植物,夏季几乎遍布于河道的滨水浅滩,第2种组的碱蓬、碱莞、苣荬菜主要分布于潜流湿地等土壤含盐量较高的区域。除此之外,可达性较强的区域距离水边和河道较远,部分植物受人为活动的影响强烈,如不定期引种、除草、修剪等出于景观构建、美化等原因的人为干扰,对种间关系构建和群落稳定性造成一定影响。第4种组与第5种组部分植物表现出明显的集群生长,如DCA排序中,速生杨出现在排序图中央,表现出群落优势性,实际其主要出现湿地周边人工林; 国槐-速生杨-兰花鼠尾草等种对,种间关系不显着,且联结系数较小,但实际却表现出相似的生态适应性,分布广泛,数量多,多见共生或伴生生长。
生态种组的划分以Spearman秩相关分析的结果为主要依据,两两之间相关性较高且显着性较强的物种多为正关联,人工干扰种对多表现出无连接或负联结。因此,为了更准确的划分生态种组,按分析方法提出了种组划分的参考依据( 表2) ,再结合物种实际的生态习性和分布生境。如第1组团中的芦苇,Spearman秩相关分析中相关性位于(0. 4 - 0. 8) 第二区间,但其生于浅滩或浅水区,χ2检验为极显着正相关,因此,应属于湿生植物组团。研究显示,正关联种对表现出较好的显着性、Spearman秩相关系数等级也较高,原始生物种之间多成正联结,多分布在靠前列的生态组团。
杜道林等[21]认为,群落成熟度愈高,物种间应存在较强的正关联,以求多物种稳定的共存。李育中等[22]认为随着演替的发展,各组成种群分别占据,进一步分化了的生态位并能和谐共生于群落之中,种对间联结关系也变得较为松散,因此演替成熟阶段的植物群落很可能表征为优势种间没有正联结,少数发生负联结,多数为单独分布。通过静态地对群落内物种间的关联性研究就可推测群落的演替动态和趋势,其结果有助于正确认识群落的基本结构与本质特征,并为植物生态环境的保护和优化利用物种资源等提供依据[23]; 但种间联结方法有着自身的缺陷[24 - 25].
湿地恢复受人工干扰较大,植物群落结构也会因环境因子的不同作用而发生变化。随着演替的进行,湿生植物的生长环境可达度小,水分充沛,如酸模叶蓼、稗、扁秆藨草等物种的数量和范围可能会扩大,如芦苇是密集型地下芽克隆植物,适应湿地水文条件,受湿地水文的限制小,在竞争中占明显优势[26].中生或旱生的物种如碱蓬、苣荬菜等范围也将增加并将在盐碱湿地中占据优势,绿地组团和人工林作为水域湿地的屏障,丰富景观效果,创造园林意境,增加乡土野趣,为湿地实现合理的植物种类选择、科学的植物群落配置与完善的湿地生态系统功能上提供了参考性的建议。湿地恢复的关键之一是湿地植被的恢复和重建。湿地植被的生长对湿地水文资源也有非常大的影响,由于植被的养分吸收作用,会直接影响到土壤相关盐分的分布及浓度。特别对于天津临港湿地,以围海造陆为起点,受到人类活动的持续影响,其湿地恢复演替的过程、植被的分布及种间关系有一定的特殊性。因此,针对湿地的不同环境条件,调整恢复方案与植物保护策略至关重要。
5结论
(1) 文中研究了滨海盐碱区植被恢复后28个主要物种间的种间关联和相关关系,多物种间总体上存在显着正关联,χ2检验结果,负关联种对较多,总体趋于独立和随机分布; 但显着种对中正负关联种对比为1. 18,且显着正关联的极显着率达到85% .
(2)OI指数多集中于0. 2 < OI≤0. 4和0. 8 < OI≤1两个区间,且基本为χ2检验正相关种对,显着性较大的OI指数也较高;DCA排序图结果与OI值基本吻合,种间距离和排布位置较好地表征了物种之间相互关系。
(3) 根据Spearman秩相关性检验,结合χ2检验与DCA分析,将28个主要物种划分为5个生态种组,分别为湿生植物种组、滨水盐生植物种组、群落演替伴生种组、过渡带物种组和受人工干扰较大的旱生植物种组。
(4)χ2检验和种间联结系数、DCA排序和Spearman秩相关性检验结果都表明,群落中多数种对间关联程度未达到显着水平,正负关联种对数量差异较大,说明该群落处于演替前期,多数物种间关系不紧密。
参考文献
[1]李 博。生态学[M].北京高等教育出版社,2000.
[2]朱圣潮。中华水韭松阳居群的群落结构与种间联结性研究[J].生物多样性,2006,14(3) :258 - 264.
[3]奇 凯,张春雨,等。赤峰市沙地油松林草本植物多样性及种间关联动态[J].生态学报,2010,30(18) :5108 - 5112.
[4]李 刚,朱志红,等。子午岭乔木群落演替过程中种间联结性分析[J].东北林业大学报,2008,36(11) :25 - 28.
[5]刘萍萍,程积民。植物种间联结关系的研究[J].水土保持研究,2000,7(2) :179 - 184.
[6]李兰兰,许 诺。七里海湿地植物种间关系的数量分析[J].水土保持通报,2014,34(4) :70 - 75.
[7]房 飞,胡玉昆,等。高寒草原植物群落种间关系的数量分析[J].生态学报,2012,32(6) :1898 - 1907.
[8]蔺雨阳,王得祥,等。秦岭中段天然油松林生态种对和功能群划分[J].东北林业大学学报,2009,37(11) :46 - 51.
[9]祁 永,李洪远。京津冀典型生态恢复案例[M].天津: 天津科学技术出版社,2014.
[10]Schluter D A. Variance test for detecting species associations with some example applications[J]. Ecology,1984,65:998 - 1005.
[11]张金屯。植被数量生态学方法[M].北京: 中国科学技术出版社,1999:79 - 87.
[12]王伯荪,彭少麟。南亚热带常绿阔叶林种间联结计测技术的研究。 IV.种间联结计测公式的探讨和修订[J].植物生态学与地植物学丛刊,1985,9(4) :274 - 285.
[13]王 琳,张金屯。历山山地草甸优势种的种间关联和相关分析[J].西北植物学报,2004,24(8) :1435 - 1440.
[14]陈圣宾,欧阳志云,等。 Beta多样性研究进展[J].生物多样性,2010,18(4) :323 - 335.
[15]Greig S P. Quantitative Plant Ecology[M]. 3rd ed. Blackwell:Scientific Publications. 1983.
[16]尹林克,李 涛。塔里木河中下游地区荒漠河岸林群落种间关系分析[J].植物生态学报,2005,29(2) :226 - 234.
[17]张 峰,张金屯,上官铁梁。历山自然保护区猪尾沟森林群落植物多样性研究[J].植物生态学报,2005,29( 增刊) :52 - 56.
[18]闫美芳,上官铁梁,等。五台山蓝花棘豆群落优势种群的种间关系分析[J].生态学杂志,2006,25(4) :354 - 358.
[19]李秋玲,范庆安,等。山西黄河湿地植被优势种群种间关系[J].生态学报,2007,26(10) :1516 - 1520.
[20]王 烨,金 山,等。浊漳河干流湿地草本植物群落优势种种间关系及功能群划分[J].生态学杂志,2015,34(8) :2109 - 2114.
[21]杜道林,刘玉成,等。缙云山亚热带栲树林优势种群间联结性研究[J].植物生态学报,1995,19(2) :148 - 157.
[22]娄彦景,赵魁义。洪河自然保护区湿地主要植被类型物种多样性与表层土壤性质的相关性研究[J].湿地科学,2008,6(1) :45 - 50.
[23]刘珏宏,高慧,等。内蒙古锡林郭勒草原大针茅-克氏针茅群落的种间关联特征分析[J].植物生态学报,2010,34(9) :1016 - 1024.
[24]郭志华,卓正大,等。庐山常绿阔叶,落叶阔叶混交林乔木种群种间联结性研究[J].植物生态学报,1997,21(5) :424 - 432.
[25]林勇明,吴承祯,等。长苞铁杉林乔木层优势种群种间关联及尺度效应研究[J].广西植物,2005,25(6) :526 - 532.
[26]付为国,李萍萍,等。北固山湿地植物群落特征及其物种多样性研究[J].湿地科学,2006,4(1) :42 - 47.