
����0������
�������о����塿������CapsicumannuumL.���Ƕ�����ʩ�������Ҫ�߲�֮һ����ʩ���γɵĵ��¡����⼰�������ջ���ʩ��������ƫʩ���ʡ�������ʢ������ˮ���ܵȣ��澳������Ӱ��ֲ�������Ĺ�����á���ˣ���չ���¡����⼰��в�������澳������ҶƬ�Ĺ�����Ա仯�о����ɷḻ���������澳������ۡ���ǰ���о���չ��Ŀǰ�йص����˺���λ���д����飬�������в��ʱ������Ļ��������кܴ��ϵ������ǿ����еȹ�ǿв���£���ϵͳ��PS��Ӧ���ĵ�DI���ױ���Ϊ�ǹ����Ƶij�ʼ��λ������������ֲ�������Ϊ�ⱥ������ʺ�CO2����������½���PS���ϵ��Ӵ�������Ч�ʺ�����ת��Ч�ʽ��ͣ����չ�ǿ����PS��⻯ѧ��Ӧ�IJ��ּ��٣������Ⱥ�ɢ�ͷ�Ӧ����������ɢ������������ϵͳ��PS�Ե�������в�Ƚ�PS������У����ƹϵ��ڵ��������¹���½�����Ҫԭ����PS���������ơ����²���������Ҷ��������н��й�����ð���Ӧø�Ļ��ԣ�Ҳ����������Ĥ�鵼�ĹⷴӦø���ԵĽ��ͣ��ҹⷴӦø���ԶԵ����˺������У�ʹ������Ĥ��PS��Ĺ��ܴ���Ч�ʺ���ת��Ч�ʽ��ͣ��Ӷ�����CO2ͬ���������͡�Kaniuga�ȱ��������µ���Ҷ�����ϣ����Ӧ���Խ��ͣ�ʹ������Ĥ��PS��ĵ��Ӵ������֣����Ʋ�λ��Ҫ���������ࡣ�����������ľ�������ʣ�Pn�����������ʣ�Tr�����ⲹ���㣨LCP����CO2�����㣨CCP�����Ȼ�Ч�ʣ�CE���½�����������Ч�ʣ�AQY����������Pn�½��Ƿ��������ƵĽ�������Ź���ǿ�ȵļ���������ҶƬChla��Chlb��Chla+b�ĺ������ӣ�Chla/b��ֵ�½���Ҷ����a�������壨CPI���������ͣ�����Ҷ����a/b�������壨LHCP���������ߡ�������ɻƹ�Ҷ���巢���������������ң����ṹ��ƻ�����Ҷ�����������٣�Ҷ���صĽ���Ӿ磬Ҷ���غ������͡���в����ֲ���������½���ԭ��һ����Ϊ����в������ģ�����в������ˮ�Ƽ������Ƚ��ͣ�����CO2�����ϻ������Ӷ����ƹ�����ã�ͬʱҲ���ڻ������˺�����в���£�ҶƬNa+�������ߣ��������ײ��ֹرգ������Ƚ��ͣ�����CO2��Ҷ��������ͣ�ʹҶ������CO2ͬ�����裬���Թ�ϵ��Ӵ���Ӱ����Խ�С�����������Ӵ��ݵ�O2�������γɣ�����H2O2�����Ҷ������H2O2�Ļ��ۣ��Ӷ��ƻ�Ҷ����Ĥ�ṹ�����¹�Ϲ����½���
������������ʹ��Ҷ�����Բ������Ƭ��Ķ���ṹ���壬���ֻ����ܽ⣬������Ŀ���١�ƫ��������[19��/12��(��/ҹ)��90?mol·m-2·s-1]���ٽ��������[15��/8�棨��/ҹ����90?mol·m-2·s-1]�£����������LCP���ⱥ�͵㣨LSP����AQY�½���CCP���ߣ�CO2���͵㣨CSP����CO2����ʱ�Ĺ�������Լ�CE�½����¶Ȳ����㣨TCP�����ͣ������������ʱ�䣨STP���ӳ���ƫ���������£������Թ���CO2����������������Ч�ʽϸߣ������������ʱ��϶̡������о�����㡿Ŀǰ����о���༯���ڵ��¡����⡢��в�ȵ�һ�澳���������˫���澳�ȶ����������ѡ��ƹϡ����ӵȹ�����Ե�Ӱ�죬��������ʩ�ڵ��͵ĵ��¡����⼰��в�������澳���߲�ֲ�������Ե�Ӱ�컹δ���������������Ĺؼ����⡿������ģ����¡����⼰��в�������澳��̽�������澳������ҶƬ������Ե���Ӧ���ƣ�Ϊ������ʩ�����е��¡��⡢�ʹ����ṩ�������ݡ�
����1�������뷽��
����1.1�������
��������������CapsicumannuumL.��Ʒ��Ϊ‘�н�4��’��������Ʒ�֣������й�ũҵ��ѧԺ�߲˻����о����������ֿ������ṩ��
����������2007�����й�ũҵ��ѧԺ�߲˻����о������������ڽ��С����Ӿ�5%NaClO����15min�����4h��Ȼ��������˫����ֽ��Φ15�������ڣ����������䣨28�棩�д�ѿ��ѡ��ѿ����һ�µ����Ӳ�������Ӫ������8cm×8cm���У�����2�s1��v/v���IJ�̿����ʯ���ϻ��ʣ�ÿ�������ʼ�5kg�����1.5kg���Ϸʣ�N-P2O5-K2O=15-15-15����
����1.2�������
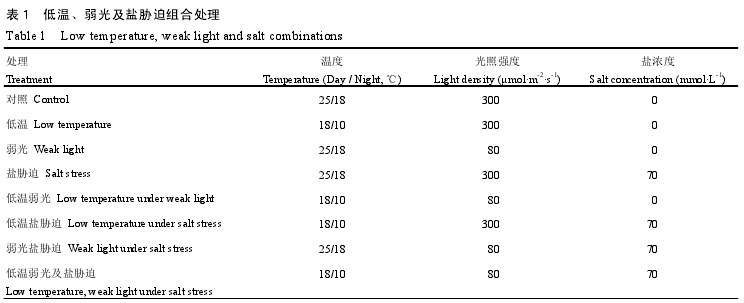
�����������5Ƭ��Ҷչ��ʱ�����˹������䣬����18��/10�棨��/ҹ�����£�80?mol·m-2·s-1�����70mmol·L-1�εĵ�һ��˫�غ�����в�ȴ��������¡����⡢��в��3���澳ͬʱ���У������¡����⼰��в����ϴ�������1�����У���в������1/2Ũ��HoaglandӪ��Һ��ECֵΪ1.71mS·cm-1���м���NaClʹ��Ũ�ȴﵽ70mmol·L-1��ECֵΪ7.50mS·cm-1����ÿ��Ӿ����1.5cm�����ʽ���120mL��Ϊ��в�ȴ���������ÿ�����1/2Ũ��HoaglandӪ��Һ120mL��δ����NaCl����������Ϊ8h/16h���������ʪ�ȱ���70%—80%�����������ƣ�3���ظ���ÿ����С��15�꣬����15d��
����
����1.3�ⶨ����
����1.3.1Ҷ���غ��������������ķ������иĽ���ѡ���������2Ƭ��ȫչ��Ҷ���ô������ȡΦ0.4cm��ҶԲƬ0.1g��������Ҵ�����ͪ���Һ10ml���ݣ����ñ���Ĥ��ڷ�ֹ��ȡҺ�ӷ�����������֯�����UV-1700����/�ɼ���ֹ��ȼƲⶨ��663��645nm���������ֵ��OD663��OD645����Ȼ��ʽ����Ҷ���غ�����
����1.3.2������彻������ѡ���������2Ƭ��ȫչ��Ҷ��Ӧ��LI-6400��Яʽ����DzⶨҶƬ��������ʣ�Pn���������ȣ�Gs��������CO2Ũ�ȣ�Ci�����������ʣ�Tr���ȡ�Ҷ�ң�2cm×3cm�����趨�¶�26�棬��ǿΪ600?mol·m-2·s-1��ÿ������ѡ3�꣬ÿ�����5�Ρ�
�����������ԡ�
����1.3.3Ҷ����ӫ���������LI-6400��Яʽ����ǵ�6400-40ӫ��Ҷ�Ҳⶨ��ҶƬ����Ӧ20min������ԭ����ӫ��������رչ⻯ѧ���Զ��⣬������ⶨ��ʼӫ�⣨Fo�����ٴ�һ�����������ⶨ���ӫ�⣨Fm����Ȼ��⻯ѧ�⣨��ǿ600?mol·m-2·s-1������ҶƬ������̬ӫ�⣨Fs�����ֺ��ٴ�һ�����������ⶨ�������ӫ�⣨Fm’�������رչ⻯ѧ�⣬��һ��Զ�����ⶨ������Сӫ�⣨Fo’����������ɱ�ӫ�⣨Fv������ϵͳ��PS�����⻯ѧЧ�ʣ�Fv/Fm����ʵ�ʹ⻯ѧЧ�ʣ�ΦPS���⻯ѧ���qP���Ȳ���������Fv=Fm-Fo��ΦPS��=��Fm’-Fs��/Fm’��Fv’=Fm’-Fo’��qP=��Fm’-Fs��/��Fm’-Fo’��������ת��Ч�ʣ�Fv’/Fm’��=��Fm’-Fo’��/Fm’���⻯ѧ��Ӧ���ʣ�Prate��=�ۣ�Fm’-Fs��/Fm’��×ҶƬ���յ�PFD�������Ⱥ�ɢ���ʣ�Drate��=��1-Fv’/Fm’��×ҶƬ���յ�PFD���ֱ��ڴ�����1��5��9��15d��ѡ���������2Ƭ��ȫչ��Ҷ���вⶨ��
����1.3.4��ָ̬�괦����15d����Ӫ����������ȡ����������ֲ�꣬��ˮ��ϴ�ɾ���������Ⱥ��¹ķ�����䣬105��ɱ��30min��80���������غ�Լ��4h���������ϲ��������ء�
����1.4ͳ�Ʒ���
�����������ݾ�����SAS����Duncan’s�¸�������ж��رȽϼ����������Է���������ͼ�����ݾ�Ϊ3���ظ���ƽ��ֵ��
����2�����
����2.1���¡����⼰��в��������ҶƬҶ���غ����ı仯
�����ӱ�2��֪���������ȣ����⡢�������в�ȴ������������ҶƬChla��Chlb��Chl��a+b��������������Chla/b��ֵ�����Ŵ���ʱ����ӳ���Chla��Chlb��Chl��a+b�����������������ƣ�Chla/b��ֵ���½����ơ���������в�ȴ�����1��5��9��15d��Chla�����ֱ�ȶ���������1.99%��11.51%��22.18%��35.45%��Chla/b��ֵ�ֱ���0.71%��1.49%��4.38%��6.88%�������������෴�ı仯���ƣ������Ŵ���ʱ����ӳ�����Chla��Chlb��Chl��a+b��������������������ǿ��������в�ȴ��������������������15d��Chla��Chlb��Chl��a��b�������ֱ�ȶ��ս�����22.16%��33.84%��25.16%��
����2.2���¡����⼰��в��������ҶƬ������ò����ı仯
�����������ȣ����¡����⡢��в�ȵ�һ�������澳������������ҶƬ��Pn���Ҹ����澳������Ч�����ڵ�һ�澳����3�����紦����9d����һ�澳��Pn�ȶ��ս�����42.84%—50.09%�������澳��Pn������56.92%—65.24%���������⡢������в�ȡ��������⼰��в�ȴ���ʱ����ӳ���Pn���Ƚ��ͺ����ߵ����ƣ��ֱ��ڴ�����5��9��9d�����ֵ�������澳��Pn����ͬ�ı仯���ƣ����洦��ʱ����ӳ���Pn���������ơ�
�������Ŵ���ʱ����ӳ�����һ�������澳��Gs�������͵����ơ����⡢������в�ȴ�����Gs���ͷ��Ƚ�С��������в�ȴ�����Gs���ͷ��Ƚϴ�����15d�ֱ�ȶ��ս�����44.83%��61.24%��83.69%��Tr����ͬ�ı仯���ơ�
�������¡����⡢�������������������ҶƬ����CO2Ũ�ȣ������������˰���CO2Ũ�ȡ����Ŵ���ʱ����ӳ���������İ���CO2Ũ�ȳ������ߺ͵ı仯���ƣ����¡�����������İ���CO2Ũ���洦��ʱ����ӳ��������ߵ����ơ����Ŵ���ʱ����ӳ�����в�ȡ�������в�ȡ��������⼰��в�ȴ����İ���CO2Ũ�ȳ��Ƚ��ͺ����ߵı仯���ơ�
����2.3���¡����⼰��в�ȶ�����ҶƬҶ����ӫ�������Ӱ��
�����������ȣ���һ�������澳������ҶƬPS�����⻯ѧЧ�ʣ�Fv/Fm�����������ͣ����洦��ʱ����ӳ���Fv/Fm�ʳ����½����ƣ���4�������⡢������в�ȴ�����Fv/Fm���ͷ��Ƚ�С��������в�ȴ�����Fv/FmӰ���������1��5��9��15d���ֱ�ȶ��ս�����3.78%��5.98%��9.87%��16.69%��PS���ϵ��Ӵ�������Ч�ʣ�ΦPS�����Ƶı仯���ơ�
�����������ȣ���һ�������澳������������ҶƬPrate�������Drate�����⡢�������в�ȴ�����Prate���ͷ��Ƚ�С��������1��5��9��15d���ֱ�ȶ��ս�����6.22%—9.94%��11.39%—19.48%��16.37%—24.17%��24.23%—26.63%��������в�ȴ�����PrateӰ���������15d�ȶ��ս�����45.44%�����⡢�������в�ȴ�����Drate���߷��Ƚ�С��������в�ȴ�����Drate���߷����������15d�ȶ���������46.33%��
�����ӱ�5��֪���������������µ������������չ�������42.2%—43.3%���ڹ⻯ѧ��Ӧ��39.0%—39.5%�������ߺ�ɢ��17.4%—18.3%ΪPS��Ӧ���ĵĹ�ʣ���ܡ��������ȣ���һ�������澳��������������������ߺ�ɢ��ʣ���ܱ��ʡ����У�������в�ȴ��������ߺ�ɢ������Ϊ���ԣ�������15d�����������57.8%���Էǹ⻯ѧ��Ӧ����ʽ�ڷ�Ӧ���ĺ�ɢ�IJ������ӽ�Ϊ���Ե��ǵ��´�����������9d�����������21.6%��
����2.4���¡����⼰��в�ȶ�������������ʻ��۵�Ӱ��
�����������ȣ���һ�������澳����������������ĸ����ʻ��ۣ��Ҹ����澳������Ч�����ڵ�һ�澳����6�����絥һ�澳�ĸ����ء����ϲ����طֱ�ȶ��ս�����35.79%—47.61%��22.53%—31.34%�������澳�ĸ����ء����ϲ����ؽ�����55.69%—65.31%��33.23%—49.59%�����������������ĸ����ʻ���Ӱ���С��������в�ȡ��������⼰��в�ȴ�����������������ʻ��۵��������ýϴ��ܸ��طֱ�ȶ��ս�����26.26%��45.48%��52.59%��
����3������
����Ҷ�����ǹ�����õ����ʻ������ǹ�����õĹ���������Ҷ���ذ���Chla��Chlb���࣬���о���Chla���Ӻ�ȫ��Chlb������Ҫ����ת�ݹ��ܵ����ã���������״̬��Chla�����н�����ת��Ϊ���ܵ����á���ͬ�澳��ֲ��Ҷ���غ���Ӱ���������졣���´��������������ƹϵ�Ҷ���غ����½������²���Ӱ��Ҷ���صĺϳɣ������Ҷ���ؽ���Ӿ磬һ����Ϊ������Ҷ���غ������Ͳ�����Ϊ�ϳ�ϵͳ���裬��������ԭ��Ҷ�����ܵ��ƻ����¡�
������������������ҶƬҶ���غ����������˻ƹ�ҶƬҶ���غ���������������ڹ���ǿ�Ⱥʹ���ʱ�䲻ͬ����,Ҳ����������ͳ��������ķ�Ӧ���ڲ����йء���в�ȡ�������������»ƹϡ�����Ҷ���غ����Ľ��͡����⡢������в�ȴ������������ҶƬChla��Chlb��Chl��a+b��������������Chla/b��ֵ�����Ŵ���ʱ����ӳ���Chla��Chlb��Chl��a+b�����������������ƣ�Chla/b��ֵ���½����ƣ���2����˵�������������£�Ҷ���أ��ر���Chlb���������ӿ�����һ�ֱ����Է�Ӧ�����Ӳ���ɫ�ظ�����������ɫ�صı������Ը���Ч�ز������Ĺ��ܣ�����Ϊв�Ⱥ��ڣ�������9—15d������ҶƬPn���������ƣ���3���������������෴�ı仯���ƣ�������в�ȴ�����Chla��Chlb��Chl��a+b��������ͣ�������15d�ֱ�ȶ��ս�����22.16%��33.84%��25.16%�����������ֲ�������������Ҫ��������֮һ��
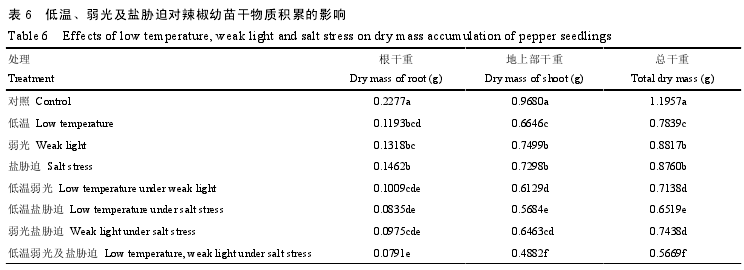
�������澳��������������ҶƬPn���½�����3�����Ӷ�ʹ��������ʻ��ۼ��٣���6��������Pn���ͼ�����������Ҳ�з��������ء����¡����������������������Pn�½���ͬʱ��Ci������˵��Pn���½�������ΪGs�½�ʹCO2��Ӧ�������£��������ڷ��������أ�Ҷ��ϸ��ʹ������ɢ���迹���ӡ�CO2�ܽ���½���Rubiscoø��CO2���������͡�RuBP���������½����ϻ����ؼ��ɷֵ��ȶ������澳в�ȶ����͵ȣ��谭��CO2�����ã��Ӷ����ϸ����϶CO2���ۣ������������Pn�½�ͬʱ��Ci���������½���˵��Pn�½��ɷ�����������ת��Ϊ��������ռ������λ����в�ȡ�������в�ȡ��������⼰��в�ȴ�����������Pn�½���ͬʱ��Ci�Ƚ��ͺ����ߣ�����Pn�½�������������ת��Ϊ����������ռ������λ��������в�ȴ�����������Pn�½���ͬʱ��Ci�½�������Pn�½�����Ҫ���������ء��������ȣ������������������ϲ������ʻ����½�������Խϵͣ�ԭ�������ֲ��Ϊ���ֲ����ղ��㣬�������������IJ����Ի�ȡ�㹻�Ĺ��ܣ���ʹֲ�����ڵĹ�ϲ��P���ָ�������ڵ��ϲ��������෴����в�ȴ���������������²������ʻ����½����Ƚϵͣ�ԭ�����������в���£�Ϊ����һ���ĸ�ϵ������ֲ��ֻ��ͨ�����Ӹ�ϵ���ܵ�λ���ֲ�����ϲ�����Խ϶�ط��䵽��ϵ��
�����������Ǹߵ�ֲ�������ù������ձ���ڵ�������������ʱ����Ĺ��ܲ���ʹFv/Fm�½������Ҷ�ҶƬ�Ĺ�ϻ�����ɲ�ͬ�̶ȵ��˺���Fv/Fm����Ϊ�Ƿ�ӳ�����Ƴ̶ȵĿɿ�ָ�꣬��ʾPS��ԭ�����ܵ�ת��Ч�ʡ���㶫���о����������������£��ƹ�ҶƬ��PS��Fv/Fm�仯����˵���ʵ��������������PS��Ĺ⻯ѧЧ�ʣ���������ơ���в�ȴ������Խ����˻ƹ�ҶƬΦPS��Fv/FmӰ�첻��
�����λ��еȶԵ��������·��ѵ��о�����������2��Ʒ�ֵ�Fv/Fm����ͬ�̶ȵ����ӣ�������������û��ʹ���ѵ�PS��⻯ѧ�����ܵ����ƣ������Dz���Ʒ�ֶԵ���������һ������Ӧ������������в�ȴ���������������Fv/Fm�ϴ���ȵ��½��������������ر������⣩��Fv/Fm��Ӱ���С��˵��������в�ȵ��µĹ����ƽ����أ�����˹�ϻ��������ˣ�����������Fv/Fm���½�������һ�ֹⱣ�����ƣ�����������Ĺ�������½�����������ڰ���Ӧ���½�����ġ�
�������澳�������������ߺ�ɢ��ʣ���ܱ��ʵ����ӣ���5�������У�������в�ȡ��������⼰��в�ȴ��������ߺ�ɢ������Ϊ���ԣ�������15d�����������57.8%��55.0%���Էǹ⻯ѧ��Ӧ����ʽ�ڷ�Ӧ���ĺ�ɢ�IJ������ӽ�Ϊ���Ե��ǵ��´�����������9d�����������21.6%�����ֱ仯��ַ�ӳ��ֲ����澳��һ����Ӧ�����������������Ⱥ�ɢ���ӿɼ���PS��͵��Ӵ������Ĺ��ֻ�ԭ����Ҷ���ص�����״����Ҷ���ؿ�Ĵ�С��أ���ֲ���ֹ��ʣ���ܵ��¹�ϻ������ƻ��ı����Ի��ơ�
������һ�ٽ���£�15��/8�棩�£��ƹ�ҶƬҶ���غ�����Ҷ����ӫ�����Ӳ����������������ͣ��ٽ�������⣨300?mol·m-2·s-1���£�Ҷ����ӫ�����Ӳ���������ߺ͵ı仯���ƣ�Ҷ���غ��������ٶȱ�����RuBp�Ȼ�ø�����������ͺ����л�����
��������÷�������½������գ�18��/10�棬270?mol·m-2·s-1���»ƹ��ϻ����ͬ������������������������������У�������һ���̶�����������Ҷ���ء�����������ø���ԵĽ��ͣ��������������������Ĺ��������˵��������һ���̶��Ͻ����˵��µ����ã�ʹҶ���غ����½��ٶȱ�����Ҷ����ӫ�����Ӳ������ߣ��Ӷ�ʹҶ���ص��Ա�����ʹ��ϵͳ��������ߡ���������������������Ҷ���غ������ͼ�Pn��Fv/Fm��ΦPS���½��ȷ�������˵�����в�ȸ����澳�����ã������������Ե�����в�ȸ����澳�������ԡ�
����4������
�������¡����⼰��в�������澳������ҶƬ������Ե���Ӧ�����뵥һ��˫���澳���������졣���¡��������в�ȵ�һ��˫�ػ������澳������ҶƬ��������ʾ����ͣ����澳��������ҶƬ����������½������������Dz�ͬ�ġ�������в�ȶ�����ҶƬ��ϲ�����Ҷ����ӫ�����Ӱ��������Ϊ�������⼰��в�ȣ�Ӱ���С�����������
�����ο����ף�
����[1]�Ź���,���̻�.�����������������������������������ʱ���Ӱ��.����ֲ��ѧ��,2006,26(9):1770-1775.





