
��������ȫ�������Ǩ���ɺ�����Ƶ��,ˮ�ֶ�ȱ�ѳ�Ϊ����ֲ�������õ���Ҫ��������֮һ[1,2],���о�������ֲ���ܵ�ˮ��в��ʱ,��������( ���ر����� CO2��Ӧ����) ������������( Ҷ��ϸ����ϻ����½�) �Լ������������ͬ���ص���ֲ��ľ���������½�[3 -5],Ҳ���о���������ˮ��в�ȿɵ���Ҷ�����ϻ������ƻ�[6],PS����������������,PS��ɫ�ص����������ɳɷֵı仯,���CO2ͬ��Ч�ʽ��ͼ�Ҷ����ӫ��仯[7,8],���ֲ��ҶƬ�Ĺ��ӫ�����Կ�����һ���̶��Ϸ�ӳֲ��Ըɺ��澳����Ӧ.ˮ��( ����������ˮ��) ��ֲ������������ڡ���ϵ��������̾������Ե�Ӱ��[1,9],��ֲ�����������̶���������ȱˮ��һ������Ӧ�Ժ͵ֿ���,ֲ��Ĺ�����������������ˮ�ֳ���ʱ���Ծ,�������ʶȵ�ˮ�ֿ�ȱ��Χ֮�����Ծ[10,11].��������,��ͬ����ˮ��������ֲ���������̬���Ե��о��ڹ��������ܵ�����,ֲ��Ըɺ��澳����Ӧ����Ӧ�ѳ�Ϊ�о��ȵ�֮һ[1,12].
��������ľ( Pistacia weinmannifolia) Ϊ�����ơ�����ľ��,���̹�ľ��С��ľ,ż����״��Ҷ����,��СҶ4 - 9 ��,��Բ�λ���״��Բ��; ��Ȼ������,������,Ŀǰδ����" ��Ҷ" �Ͳ��溦����,��������辰����,����������̻�����.����ľ�����ϸ��Ⱥӹȷֲ��㷺[13],�õ��������ȸ���,����ϡ��,�����ɺ�ƶ�,���ҹ����ּ������ѵ���������֮һ[14].ǰ�˶�����ľ�о�����,����Ҫ�������ľ�����Ի�������[13,14],��������ľ��ˮ��в���¹��ӫ�����Ե��о����Ǻ���,�ݴ�̽������ľ������õ����ƻ�������������ˮ�ֶ�����ϵ���о���δ������.����ͨ�����Կ�ˮģ��Ƚ��о���ͬ����ˮ������������ľҶƬ�Ĺ��ӫ������,����̽������ľ��������ˮ�ֱ仯�ķ�Ӧ����Ӧ����,Ϊ����ľ��ѧ��ˮ�����Ⱥӹ������̻��ṩ��ѧ����.
����1 �����뷽��
����1. 1 ����ظſ�
���������( 102°45'E,25°04'N) λ��������ҵ��ѧУ��,���� 1946m.�õش�����������,���±仯��,��ƽ�����¶��� 20������; �ļ���������,������ռȫ�������� 60% ����,ƽ������ 22��; �ﶬ�����ճ���,�쾦����,ÿ������ƽ���� 20 ������,���� 230 Сʱ����,���� 4 ������,ȫ�½�������ռȫ���3 - 5% .��ƽ������ 15�� ,������� 2200 Сʱ,�꽵ˮ�� 1035mm,���²�Ϊȫ����С,���²�ϴ�.
����1. 2 ʵ�����
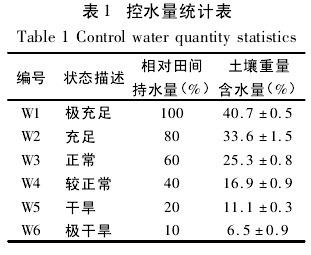
����ʵ��������Է���ֲ�����������2 ��������ľ��ľ( ��Լ70cm) Ϊ����,��������Ϊ������������,�ʵ�ճ��,�ܶ� 0. 93g·cm^- 3,����ˮ�� 40. 7%,��Ч�����ס��طֱ�Ϊ 58. 14��5. 25��55. 76mg·kg^- 1,pHֵ 4. 9.2013 �� 10 �� 18 ����������ѡ���������������С����һ�µ���������ľ 18 ��,�����ڿտ������ߺõ������,ÿ 3 ��Ϊһ��,�� 6 ��.ʵ�鹲���� 6 ����ͬ����ˮ���ݶȴ���( �� 1) : ������( W1) ������( W2) ������( W3) ��������( W4) ���ɺ�( W5) �����˸ɺ�( W6) ,�ֱ����ˮ��Ϊ��������ˮ���� 100%��80%��60%��40%��20%��10% ,�� HH2 ������ʪ�ȼƲ������������ˮ���ֱ�Ϊ 40. 7 ± 0.5% ��33. 6 ± 1. 5% ��25. 3 ± 0. 8% ��16. 9 ± 0. 9% ��11. 1 ± 0. 3% ��6. 5 ±0. 9% ,ÿ�� 18: 00 ���Ҷ�ʱ������Ӧ��Ų�ˮ,������������괦��,��ȷ����ͬ����������ˮ���ȶ�.ֲ������Ӧ��������ˮ�ִ�����( ʵ������� 1. 5 ����) ��ʼʵ��,ÿ�鴦�� 3 �� 3 ������ľ,ÿ������ľѡ�� 3 ֦Ҷ,�ⶨ���ϲ�����Ҷ����ӫ�����.
����1. 3 �ⶨ����
����1. 3. 1 ��ϲ����ⶨ
����ѡ����������( 2013 �� 12 �� 2 ��) ,�� LI -6400 ��Яʽ���ϵͳ�ⶨ��( ������Ȼ��Դ����� CO2)�Բ�ͬ����ˮ�ִ���������ľҶƬ�Ĺ�ϲ������вⶨ.����ʱѡ�� 6400 -15 СҶҶ��,�ⶨ�����Ҷ��������ľҶƬ���ռҶ�����( 0. 785cm²) �İٷֱȹ���.ÿ������ľѡ�� 3 Ƭ���վ�һ��ҶƬ�����ϼǺ�,�� 10:00 -11:00 ����.�ⶨָ���������ľҶƬ�� Pn��Tr��Gs��PAR��Ci ��,���� WUE = Pn/Tr ���п���.
����1. 3. 2 Ҷ����ӫ������ⶨ
����Ҷ����ӫ����ز����� Li -6400 ����ǵı�ӫ��Ҷ�ҽ��вⶨ.12 �� 4 �� 22:30( ҶƬ�ѽ��г�ֵİ���Ӧ) ,������ȱ�Ǻõ�ҶƬ,�ⶨ����С��ʼӫ�� Fo �����ӫ�� Fm.12 ��5 ������11:00( ����Ӧ״̬) ,���,����ǹ���ҶƬ��������ӫ��Ҷ����,�ⶨ������Сӫ�� Fo'���������ӫ��Fm'����̬ӫ�� Ft.�����¹�ʽ����:
�����⻯ѧ�������Ч��: Fv/Fm = ( Fm - Fo) /Fm
��Чӫ�����: Yield( ��Fv/Fm) = ( Fm'- Ft) /Fm'
�������Ӵ�������: ETR = Yield × PAR ×0. 84 ×0. 5
�⻯ѧ����ϵ��: qP = ( Fm' - Ft) /( Fm' - Fo')
�ǹ⻯ѧ����ϵ��: NPQ = ( Fm - Fm') /Fm' = Fm/Fm' - 1
1. 4 ���ݴ���
�������� Excel2003 �� SPSS11. 5 �������ݷ�������ͼ.���رȽϲ��� LSD ���� Sig =0. 05 ����ˮƽ�¶Բ�ͬ���ݼ�IJ�����м���Ƚ�.
����2 ��������
����2. 1 ��ͬ����ˮ�ִ���������ľҶƬ�����������������Ӱ��.
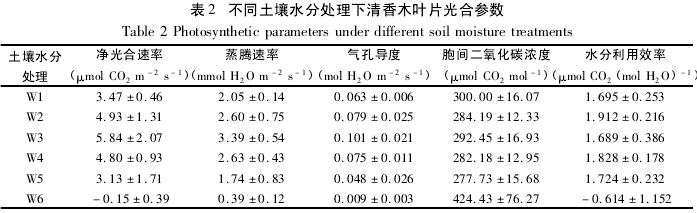
������������ʺ�����������ֲ�������������������[15],ͨ�� Pn ��ʱ Tr Ҳ�ϸ�[16].��� 2,W1 -W3,Pn �� Tr ������; W3 - W6,Pn �� Tr ��С.Pn �� Tr �ڲ�ͬ����ˮ�ִ����¸�����������һ��,W1��W2��W3��W4��W5 �������Ÿ��� W6( p <0. 01) ,W3 ���Ÿ��� W1( p <0. 05) ,W5 ���ŵ��� W2��W3��W4( p < 0. 05) ,�� W5 �� W1 �����Բ���( p > 0. 05) .WUE ֵ�� Pn �� Tr ����,���ɺ�( W6) ������WUE ֵΪ��( �� 2) ,���ŵ���ǰ���� WUE ֵ( p < 0. 05) ,�� W1 - W5 ������,WUE ֵ���в���,���֮�������Բ���( p >0. 05) .Gs ��ʾ�����ſ��ij̶�,Gs �ϴ�ʱֲ���ܹ�˳���ؽ���ˮ��������,Gs ��Сʱ����ˮ�ֵ���ʧ[17].
������� 2,����ľ Gs �� W3 ����ˮ�ִ�����ֵ���,�����Ÿ����������鴦��( p <0. 05) ,������ˮ���ϸ�ʱ( W1 - W2) ,Gs �����Ų���( p >0. 05) ,��������ˮ���ϵ�ʱ( W4 - W6) ,Gs ���ż�С,���֮���������( p <0. 05) .
������ͬ����ˮ�ִ���������ľҶƬ Ci �仯������ Pn �����෴,��� 2,Pn ��Խϴ�ʱ( W1 - W5) ,Ci�ϵ�,���� Pn ���ż�Сʱ( W6) ,Ci ��������,Ci �� W1 - W5 �֮�������Բ���,�� W6 ʱ��������( p< 0. 05) .
����2. 2 ��ͬ����ˮ�ִ���������ľҶƬҶ����ӫ�������Ӱ��
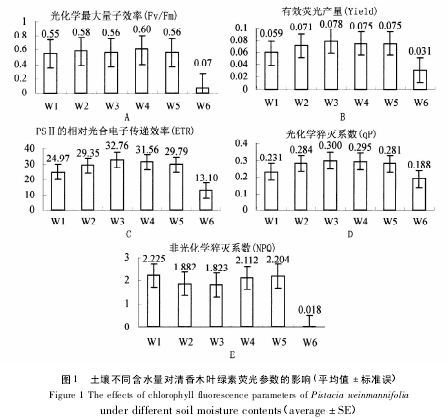
������ͼ1A,����ľҶƬ Fv/Fm ֵ�� W4 ʱ���,���� W1��W2��W3��W4��W5 �֮�������Բ���( P >0.05) ,W6 ʱ��С,��Ϊ���ֵ�� 11. 7% ,W6 ��ǰ���鴦�����켫����( P < 0. 01) ; ��ͼ 1B,W1 - W3,����ľҶƬ�� Yield ���,W3 - W6 ��С,���� W6 �����Խ���,Yield �� W1 - W5 ���α仯֮����첻����( P >0. 05) ,�� W1 �� W3 ��������( P =0. 032) ,W6 �� W1��W2��W3��W4��W5 ֮����켫����( P <0. 05) ; ��ͼ 1C,ʵ��������ľҶƬ�� ETR �� W3 ����ˮ�ִ�������´ﵽ���ֵ,W1��W2��W4��W5��W6����·ֱ�Ϊ���ֵ�� 76. 20%��89. 58%��96. 33%��90. 93%��39. 98%,W1 �� W3��W6 ��������( P < 0.05) ,�� W2��W3��W4��W5 �֮�������Բ���( P > 0. 05) ; ��ͼ 1D,��ͬ����ˮ�ִ���������ľҶƬ qPֵ��ͬ,ǰ��������ˮ�ִ����֮�������Բ���( P > 0. 05) ,W6 �� W2��W3��W4��W5 ��������( P < 0.05) ,�� W6 �� W1 �����Բ���( P = 0. 32 > 0. 05) ; ��ͼ 1E,W1 - W3,����ľҶƬ�� NPQ ֵ��С,W3- W5 ���,���֮�������Բ���( P > 0. 05) ,W6 ����ˮ�ִ����� NPQ ֵ�����½�,��ʱ PS���ܵ��ƻ�,�Ⱥ�ɢ�����½�.
����3 ����
���������о��������� Pn ���͵����ͷ������������ؿ��Ը���ֲ��ҶƬ Ci �ı仯���ж�,�� Pn �½������� Ci ����ʱ������Ϊ Pn ���½���Ҫ����������������,��� Pn �½���ͬʱ Ci �����߲���,����Ҫ�Ƿ�����������������[18,19].ʵ����,W1 - W2,Gs ��������,Pn �� Tr Ҳ������,�� Ci ������С,��ʱ��Ҫ�Ƿ�����������������ľҶƬ�������; W2 - W5,Gs��Pn��Tr �� Ci �ı仯���ƻ���һ��,����Լ������֪ Pn �� Tr �� Gs ���������,���ϵ���ֱ��0. 960 ��0. 990,��Ҫ����������Ӱ������ľҶƬ�������; W6 ʱ Pn ���ż�С,Ci ȴ��������,��ʱ������ʽ�����Ҫ�����ڷ��������������.����ɳ��ҶƬ���о����������ɺ�в�ȼӾ��������һ�ٽ��,�ٽ��֮ǰ���������������������,֮���ܷ�������������[20],���������о����һ��.
����ֲ��� WUE ��,�����̶���λ���� CO2�����ˮ��С,ֲ��Ľ�ˮ����ǿ,�ͺ�,��������[21].W1- W5,WUE �֮�������Բ���,˵����δ�ﵽ�ɺ�в����ֵ֮ǰ,����ľ���нϸߵ�ˮ������Ч��,Ciֵ�� W1 - W5 �֮�������Բ���Ҳ������ϸߵ� WUE �й�.W6 ʱ����ľҶƬ WUE ֵΪ��,ͬʱPn��Tr��Gs ֵ���Ž���,˵�� W6 �ѳ�������ľ�ɺ�в����ֵ,������ľ�ɺ�в����ֵ������������ˮ��Ϊ 11. 1%�� 6. 5%֮��.W3 - W6,WUE ��������С,���� Heitholt �ķ���һ��,�ʶȸɺ���ʹֲ��ˮ������Ч�����[22],��ɽ�ӡ��ɶ��Bľ�Ƚ϶�ֲ����о�Ҳ�����ƹ���[20].
����Ҷ����ӫ�������ֲ�������ù����й�ϵͳ�Թ��ܵ����ա����ݡ���ɢ������ȷ�����ж��ص�����,�뷴ӳ�������"������"�Ĺ�����彻��������ȸ����з�ӳ�������"������"���ص�,�κλ������ӶԹ�����õ�Ӱ�춼��ͨ��ҶƬҶ����ӫ���ѧ��ӳ����[23].Fv/Fm �ǰ���Ӧ������ PS������⻯ѧЧ��,��ӳ�� PS������������ת��Ч��,�����ȶ��� Fv/Fm ֵ�����ⷴӦϵͳδ�ܵ�����[24 -26],в�������¸ò��������½�.ʵ����,W4 ʱ����ľҶƬ�� Fv/Fm ֵ���,�� Fv/Fm ��ǰ���鴦���֮�������Բ���,˵������ľ�� W1 - W5 ˮ�ִ����²���в�Ȼ���в��,��ⷴӦϵͳ��δ�ܵ��ƻ�,�����Ÿɺ��Ӿ�( W6) ,Fv/Fm ֵ�������½�,��ʱ PS������.Yield �� ETR ���������,�仯����һ��,��������ˮ�ֺ����ɸߵ��;���������С,�� W3 ������ֵ���,�� W1 �� W3 ��������,˵��������ˮ�ֹ��ڳ����в������ľ����,��ʱ�ʵ���������ˮ�ֿ��������ľҶƬ�Ĺ������Ч��.���ɺ�( W6) �ƻ��� PS��,PS��Ӧ���Ĺ�������������,�������۵��Ӵ���Ч���½�,���� W6 ˮ�ִ����� Yield��ETR ���Ž���.
����qP �� PS������ɫ�����յĹ������ڹ⻯ѧ���Ӵ��ݵķݶ�,qP ���ͷ�ӳ PS���п��ŵķ�Ӧ���ı����Ͳ��� CO2�̶��ĵ��Ӽ���[27].NPQ ��ӳ��������ɫ�����յĹ��ܲ������ڵ��Ӵ��ݶ�������ʽ��ɢ���Ĺ��ܲ���,��ֲ�ﱣ�� PS�����Ҫ����[28].W1 - W3,����ˮ��в������,PS��Ӧ���Ŀ��ų̶ȱ��,����ɫ�����չ��ܸ�������ڹ⻯ѧ���Ӵ���,����ĵ��Ӳ��� CO2�̶�,���� qP ������,NPQ ��С,����ľ�������Ч������; W3 - W5,NPQ ������,ֲ�����ܵ��ɺ�в��,����ͨ�����ֽϸߵ��Ⱥ�ɢά�ֹ�Ͻṹ PS������,Fv/Fm ֵ�� qP ֵ��������ֵ�����Ų���Ҳ���� PS��Ӧ����δ���ƻ�,�����ɺ���( W6) ,NPQ ͻȻ���ż�С( Ϊ W5 �� 0. 83%) ,��ʱ����ľҶƬ�Ĺ�ϽṹPS�����ܵ���,�����ܵĺ�ɢ����Ҳ����ɥʧ����,����ľҶƬҲ���̿�ή.
����4 ����
����( 1) ����ˮ�ּ�����ͼ��ɺ�ʱ���������ص�������ľҶƬ������ý���,����ˮ���������ʱ����������������ľҶƬ�������.
����( 2) ����ľ�ɺ�,�����ɺ�δ�ﵽ��ֵǰ���нϸߵ�ˮ������Ч��,�ʶȸɺ������������ľ�Ĺ������Ч�ʺ�ˮ������Ч��.
����( 3) ���ɺ��ƻ�����ľҶƬ�Ĺ�Ͻṹ PS��,ʹֲ���ή����.���Ϻ�������������ľ,��ɺ�в����ֵ������������ˮ��Ϊ 11. 1 �� 6. 5%֮��.
���������
����[1]����֥,�ŵ��,����ƽ,��. ���ױ����ݵ��ԭ��ͬ����ˮ���ݶ�����ݵĹ������[J]. ��̬ѧ��,2009,29( 10) : 5271 -5279.
����[2]����,������,������. �״�Ҷ��ͬˮ��״���¹�����ʼ���Ҷ����ӫ�����Ե��о�[J]. ����ֲ��ѧ��,2005,25( 11) : 2226 -2233.
����[3]Teskey R O,Fites J A,Samuelson L J,et al. Stomatal limitations to net photosynthesis in Pinus taeda L. under different environmental conditions[J]. Tree Physiology,1986,2( 1/2/3) : 131 -142.
����[4]������,ʦ�춫,��ع��,��. ���������˹��ֶ���˹��( Populus russkii) ���½���( P. alba var. Pyramidalis) ������Է���[J]. �ɺ�����Դ�뻷��,2012,26( 9) : 67 -72.
����[5]Zhang S Y,Zhang G C,Gu S Y,et al. Critical responses of photosynthetic efficiency of goldspur apple tree to soil water variation insemiarid loesshilly area[J]. Photosynthetica,2010,48( 4) : 589 - 595.
����[6]��ѩ��,����ï. ת C4 ���ø����ˮ����ϵ�Ŀ�����������[J]. ֲ������ѧ��,2001,27( 5) : 393 -400.
����[7]Ф����. ʩˮ����ë����ɳ�� 4 ������ֲ��Ҷ����ӫ���Ӱ��[J]. �ݵ�ѧ��,2001,9( 4) : 296 -301.
����[8]Kevin O. Imaging of chlorophyll a fluorescence: theoretical and practical aspects of an emerging technique for the monitoring ofphotosynthetic per-formance[J]. Journal of Experimental Botany,2004,55: 1195 - 1205.
����[9]�ΰ���,������,�Ž���,��. ���Ⱥӹ�ֲ���ָ����ֹ����ˮ���������ԵĹ��ЧӦ[J]. �ɺ�����Դ�뻷��,2013,27( 12) : 112 -118.
����[10]Montanaro G,Dichio B,Xiloyannis C. Shade mitigates photoinhibition and enhances water use efficiency in kiwifruit under drought[J]. Photo-synthetica,2009,47( 3) : 363 - 371.
����[11]����,�ƽ���. �Ⲽ��ɳĮ�иʲݶԲ�ͬˮ�ֺ����ֹ�Ӧ�Ĺ��������Ӧ[J]. ֲ����̬ѧ��,2009,33( 6) : 1112 -1124.
����[12]ŷ����,�²�,��ѧУ. �ɺ�������������ü�����������Ե�Ӱ��[J]. ��̬ѧ��,2012,32( 8) : 2612 -2619.
����[13]����,���Ͼ�,������,��. ���ϸ��Ⱥӹ� 4 ��ֲ����������о�[J]. ������ҵ��ѧ,2006,35( 2) : 9 -16.
����[14]����ǿ,�Ծ,���Ͼ�,��. ˮ��в�ȶԻ���ľ������ľ�����Ӱ��[J]. ������ҵ��ѧѧ��,2003,25( 3) : 55 -59.
����[15]������,��ǿ,�Ժ�,��. ��ɺ�����������С������ʺ��������ʶ�������������Ӧ[J]. �ɺ�����Դ�뻷��,2014,28( 7) :56 - 61.
����[16]����,��ƽ,������,��. �µغ������Ұĵ��( Melastoma candidum) ҶƬ�Ľ����������������彻������[J]. �ȴ����ȴ�ֲ��ѧ��,2005,13( 2) : 109 - 116.
����[17]������,����,����÷,��. ������ʳ�����տ�������õĹ���Ӧ���߷���[J]. �й���������ѧ��,2013,35( 4) : 405 -410.
����[18]�����,���Ǿ�,�,��. ���������̻���é����Ҫֲ���ֹ�������о�[J]. �ɺ�����Դ�뻷��,2010,24( 7) : 138 -143.
����[19]Chartzoulakis K,Patakas A,Kofidis G,et al. Water stress affects leaf anatomy,gas exchange,water relations and growth of two avocado culti-vars[J]. Scientia Horticulturae,2002,95: 39 - 50.
����[20]���,�Ź��,������,��. �����ɺ�в�ȶ�ɳ��ҶƬ������úͿ�����ø���Ե�Ӱ��[J]. ��̬ѧ��,2013,33( 5) : 1386 -1396.
����[21]��ѧ��,��־ƽ,����,��. �ƶ���ɳ�ظ�Ҷ�ʵ� 3 ����Ҷ���ֵĹ�����Լ�ˮ������Ч��[J]. �ɺ�����Դ�뻷��,2008,22( 10) :188 - 194.
����[22]Heitholt J J. Water use efficiency and dry matter distribution in nitrogen and water - stressed winter wheat[J]. Agronomy Journal,1989,81:464 - 469.
����[23]������,�����,����,��. �����ɺ����������������ӫ�����Ե�Ӱ��[J]. ��̬ѧ��,2013,33( 13) : 3933 -3940.
����[24]Jones D T,Sah J P,Ross M S,et al. Responses of twelve tree species common in Everglades tree islands to simulated hydrologic regimes[J].Wetlands,2006,26( 3) : 830 - 844.
����[25]Lavinsky A O,De Souza Sant C,Mielke M S,et al. Effects of light availability and soil flooding on growth and photosynthetic characteristics ofGenipa Americana L. seedlings[J]. New Forests,2007,34( 1) : 41 - 50.
����[26]������,�ﳤ��. ��ʩ��˼����в�������������ϵ�����������Ե�Ӱ��[J]. �ɺ�����Դ�뻷��,2013,27( 9) : 113 -118.
����[27]����,�¿���,��ε,��. �����Ѻ����˽���ˮ���ݶȵ���Ӧ����[J]. ��̬ѧ��־,2011,30( 3) : 464 -470.
����[28]������,��ƼƼ,������,��. �������ƹ�Ѩ������������Ӧ��[J]. Ӧ����̬ѧ��,2011,22( 9) : 2343 -2347.





