����ժ Ҫ����������������˫��˾���������ѧ�о����٣�˫��˾�������ϵͳ�����ȱʧ����Ŀ�ġ�����˫��˾�����Ȼ����ϵͳ����ͼ����������ٽ�������˫��˾�����ѧ�о��е������Ӧ�á�����������������������ʼ���������ͻ����鹲���Է�����������Ŀǰ�����Ѳ����˫��˾�����Ȼ��������ϵͳ�����о����������˫��˾�������֪��Ȼ�������Ի���Ϊ6����ͬ���͵����������3�����صĸ�������������III�ͼ���VI�����ɽ�һ������Ϊ��ͬ��������Ⱥ������III������˫��˾�����Ȼ��������Ҫ���ͺ����Ƽ��塣����VI������Ա���ͷ�����ḻ������������ˮƽ�����ַ����ķ�������ȫһ�¡������ۡ�����ȫ�桢ȷ�ؽ�ʾ�����з�������֮���ϵͳ�����ϵ��������˫��˾�����Ȼ����ϵͳ�ķ��������������ϵ����Ϊ���˫��˾���Ȼ��������ͼ����ṩ��Ҫ�����۲ο��ͷ������ݡ�
�����ؼ��ʣ���˫��˾���; ��������; ������ʼ����; �����鹲����; ������;
����Abstract����[Background] In the past, there were few studies on the taxonomy of bifidobacteria plasmids, and the phylogenetic classification method of Bifidobacterium plasmid was missing. [Objective] In order to establish a method for phylogenetic classification and identification of natural plasmid in Bifidobacterium, and increase understanding and application of plasmids in the research of bifidobacteria biology, [Methods] we conducted to phylogenetic classification of all sequenced plasmids in Bifidobacterium by the analysis of phylogenetic relationship of replication initiation protein and genome colinearity. [Results] The results showed that all known natural plasmids of Bifidobacterium were divided into six different types of plasmid families and three unique complex plasmids. Plasmids of type III and VI family were further divided into different subtype groups. Plasmids of the type III and VI family can be further divided into different subtype groups. Plasmids of the type III family were the main type and dominant family of natural plasmids in Bifidobacterium. Plasmid subtypes of type VI family were most abundantly classified. At the plasmid family level, the classification results of the two methods were completely identical. [Conclusion] These results comprehensively and accurately revealed the phylogenetic classification relationship between all the plasmids that were analyzed, and established the phylogenetic classification standard and method of natural plasmid in Bifidobacterium, which provided important theoretical references and classification basis for the future classification and identification of natural plasmids in Bifidobacterium.
����Keyword����Bifidobacterium; Plasmid classification; Replication initiation protein; Genome colinearity; Replicon;
����˫��˾����������˺Ͷ��������������ֳ���Ϳ�ǻ�Ȼ����У����˺Ͷ���������Ҫ�Ĺ�������˫��˾�����ͨ�������²�����������ǿ����Ӧ����������Ѫ�����ɺ���Ӫ���ȷ�ʽ����ά�ֺ͵���������̬ƽ�⡢�ٽ������������淢���Ź㷺�һ�������������[1,2,3]��˫��˾���֤ʵ���ڼ���ʵ������ѡ���Զ�λ����ֳ�������ڰ�֢���������и߶�����Ļ����������[4,5]���о�������˫��˾����˺Ͷ�����ʳ�������ͼ����������[6,7]��Ŀǰ��˫��˾�����ʳƷ��ҽҩ��ҵ�㷺Ӧ�ã���Ʒ�����ڶ࣬������Ҫ�ľ��ü�ֵ��������塣
����Ŀǰ��˫��˾�������70����(��)�ֺ�10�����ֱ���ȷ����ͼ���(http://www.bacterio. net/bifidobacterium.html)�����������������\����Ϣ����(National Center for Biotechnology Information��NCBI)��RefSeq���ݿ�(��ֹ2019-10-10)��˫��˾�������38���������������б���¼������23���������Գ�˫��˾�(Bifidobacterium longum)��15����������˫��˾����������֡���Щ���ݱ�����˫��˾�����֪��Ȼ��������ϡ�٣���Դ�Ƚ��ѷ����������ֿ��ܲ�Я����Ȼ�����������ֲ��������Ե����������ԡ����⣬����˫��˾������о���Ҫ�漰�������롢���������幹��[8]��������˫��˾�����������о����٣�������˫��˾�“��”ˮƽ�Ͽ�չ������ϵͳ����ѧ�о���δ��������

������������һֱ��ϸ�������о��Ļ�����������⡣���������Ŵ������Էḻ�������ʸߣ�����û��һ��ͨ����ȷ�ķ������Խ�������Ȼ�������з���[9]��Ŀǰ��ʹ����㡢����ķ�������������о������������������Է��෨[10]���÷�������ͼ�������ʱ����Ҫ���з����IJ������Թ�����֤ʵ�飬��ʱ������Ч�ʵ��¡�ͬʱ���������������븴�ơ����䡢ά�ֶ������������أ����ӻ��Ƹ��Ӷ���������û�б���ȫ���������һ�������˸÷�������Ч�Ժ�ȷ�ԡ���ˣ��÷����Ѳ�������������ʱ�������²��������ķ���ͼ�������������������������[11,12]���ɳ�ø[13]��Ϊ���ӱ�ǵ�����ϵͳ��������б�����ǰ��������������������Ȼ��������ʵ���и÷���ֻ���к���(���˾���[12]�����������˾�[14]�������[15]���������[16])�еõ��˷�չ��Ӧ�ã����������������δ���������������ڰл������ƣ�ֻ�����ڿ�ת�������ķ��࣬���Ӧ�÷�Χ����[13]��
����˫��˾�����˾����������о���Ӧ�õĵ��ʹ����������ǵ���Ե��ϵ(˫��˾����ڷ��߾��ţ���˾����ں�ھ���)��Ⱦɫ��(G+C)mol%����(˫��˾�Լ60%����˾�Լ40%)�����Զ����ˣ���Ȼ�����Ķ����Ժͷ���ܿ���Ҳ���Ų�ͬ��֮ǰ����������������ʼ����(replication initiation protein��Rep)��/������ȫ���������У����ǽ��������ֲ�ͬ����Ȼ����ϵͳ����������ɹ�Ӧ������˾�����Ȼ��������[17,18,19]�����÷����ܷ���������Ե��ϵ��Զ��(G+C)mol%������˫��˾�������ϸ������������ö�֪�����������в����˫��˾���Ȼ������Ϊ�о�����ͨ������Rep�����������������������鹲���Է�����̽������˫��˾���Ȼ����ϵͳ�������������ͷ�����ϵ��Ϊ���˫��˾���Ȼ�������ࡢ������Ӧ���ṩ��Ҫ�����۲ο��ͷ������ݡ�ͬʱ�����о��ɽ�һ����������Rep�������������鹲���Է��෨�Ը�(G+C)mol%����ϸ����Ȼ�����ķ���Ч��������Ϊ�����һ��ͨ�á���Ч��ϸ����Ȼ����������ṩ���ӳ�ʵ�����ۻ�����ʵ�����ݡ�
����1 �������뷽��
����1.1 ����������������
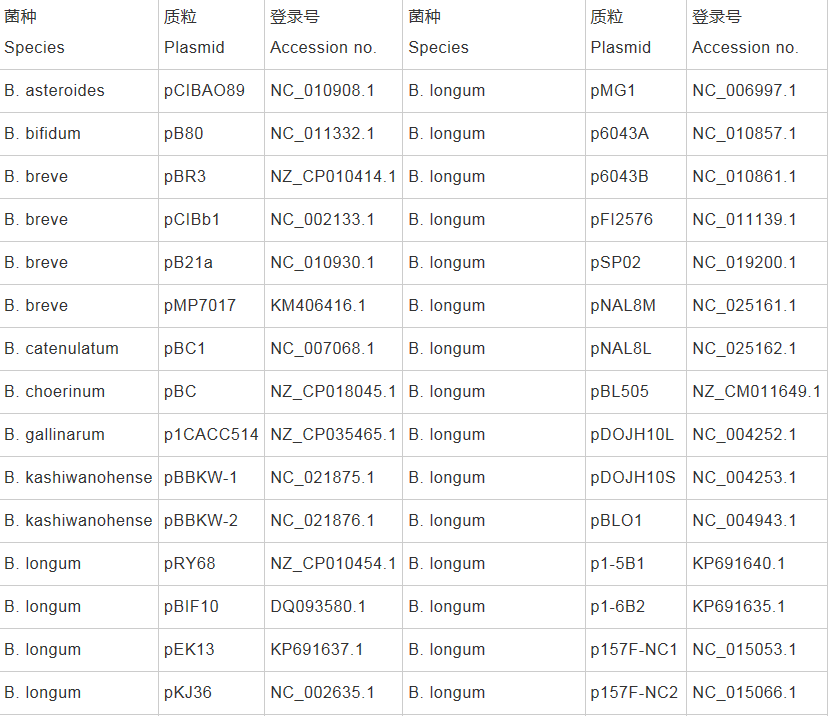
������������������������������NCBI��Nucleotide��RefSeq���ݿ⣬������ϸ��Ϣ����1��
������1 Nucleotide��RefSeq���ݿ���¼��˫��˾�����������������
����1.2 ������
����1.2.1�� �������������е��ռ��������ͳ�������
��������Nucleotide��RefSeq���ݿ���������¼��˫��˾��������������У�ɸ����������������������Rep���ױ����������ȫ��Rep�������н���BLAST���бȶԷ����������䱣�ؽṹ����ȷ�������塣����DNASTAR 7.1.0 EditSeq�����Ժ�����������н��д洢���༭�ȴ�����
����1.2.2�� ������Ͱ���������бȶԺ��ظ����з���
��������MEGA 7.0[20]������Muscle�����к������������бȶԣ��ȶԲ���ΪĬ�����á�DNA�ظ����з�������DNASTAR 7.1.0 GeneQuest�������У���С���г�������Ϊ8 bp���������ΪС��200 bp��
����1.2.3�� ����Rep����ϵͳ����������
��������MEGA 7.0������Neighbor-Joining (N-J)����������Rep����ϵͳ������������ϵͳ�������������Bootstrap������չֵ����Ϊ1 000��������������Ĭ�����á�
����1.2.4�� ���������鹲���Է���
��������Mauve 2.4.0[21]�����������������鹲���Է�����Match Seed WeightΪ13������������ΪĬ�����á������鹲���Է����У�����������Ĭ����С�������С�
����2 ����������
����2.1 ��˫��˾���Ȼ������������
������������ɸ�������ĿǰNucleotide���ݿ���¼˫��˾�����DNA����103��(����RefSeq���ݿ���¼��38��˫��˾�����DNA����)������41������Ϊ�������������С�ͳ�Ʒ������֣�˫��˾�41�����������������ܳ�387.77 kb��Я��399�����ױ������С��ΧΪ2.11-190.18 kb��ƽ��(G+C)mol%����Ϊ61.39%�������(27/41)��������B. longum�������(60/70)������δ������Ȼ����(��2)����Щ���������˫��˾������ձ��С�����������ѷ���ƽ��(G+C)mol%�����ӽ�Ⱦɫ�壬�����ֲ��������Ե����������ԣ�B. longum��˫��˾�������Ȼ������������ḻ�����֡�������BLAST�������֣�38��˫��˾�����������42��Rep���ף�����35������ֻ����1��Rep���ף�3�������������ж��Rep���ף�����3������û�б�����֪��Rep���ף�����Rep�����ݰ����������еı��ؽṹ����Գ�������Ϊ6����ͬ�ĵ����壬�ֱ��ΪRep_1��Rep_3��RepA_C��Rep*��Rep_trans��Viral_Rep (��2)�������˫��˾������(38/41)��������Rep���ף���ЩRep�������о���һ����ͬԴ�ԺͲ����ԣ����Rep������Ϊ˫��˾�����ϵͳ�����ṩ���õķ��ӱ�Ǻ����ݡ�
������2 ˫��˾��������������Rep����������
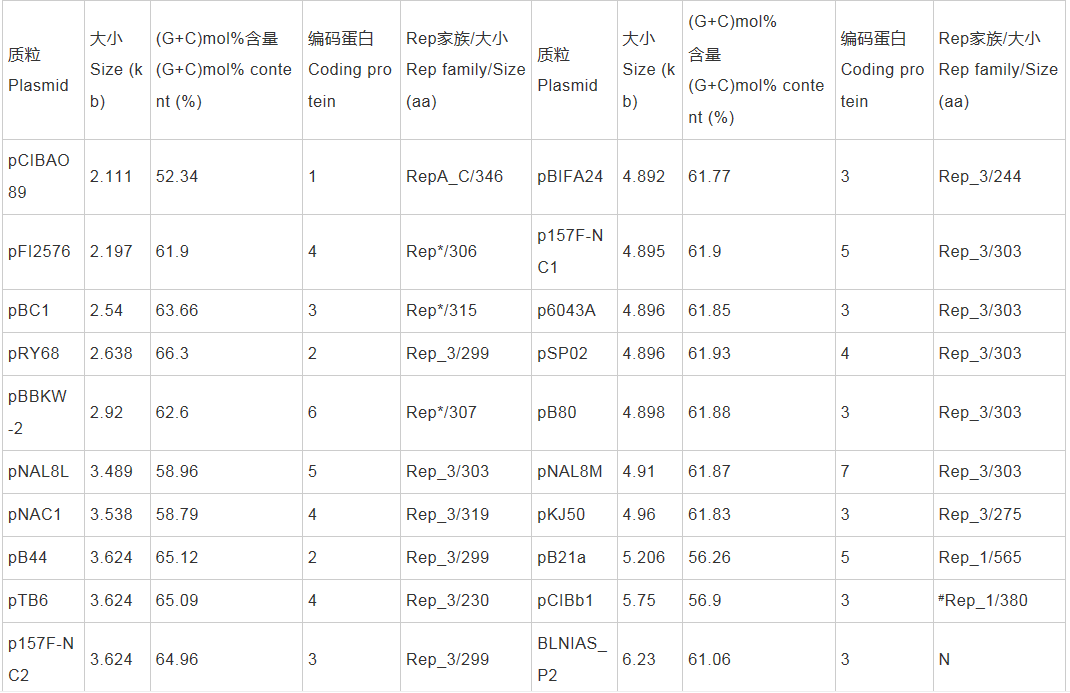
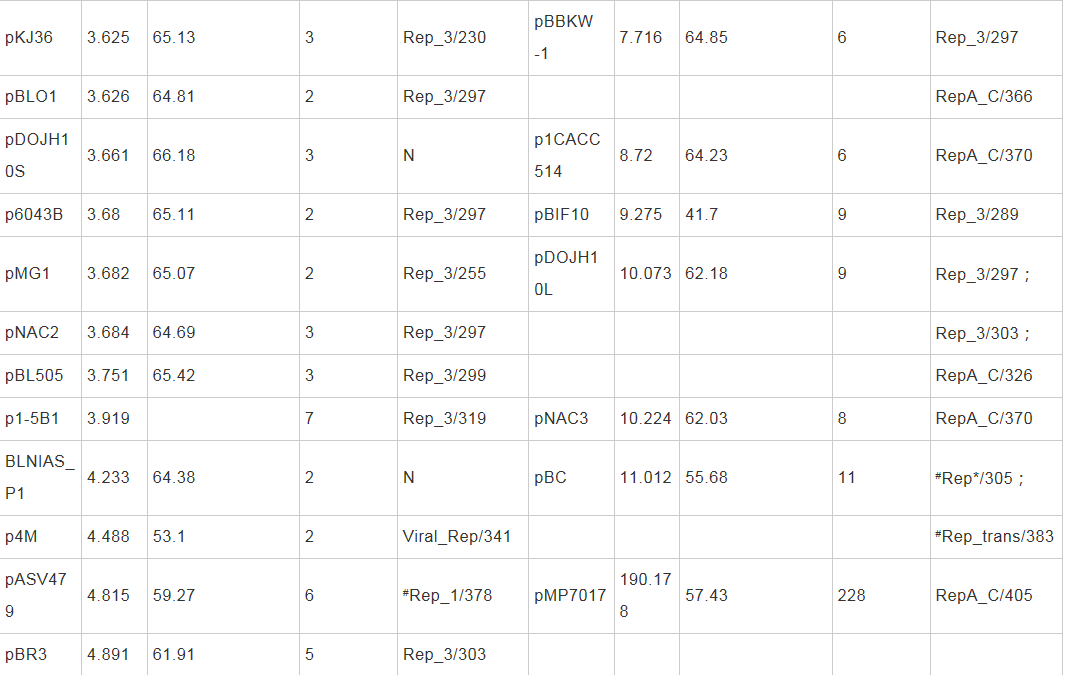
����ע��Rep_1��pfam01446��Rep_3��pfam01051��RepA_C��pfam04796��Rep*��pfam03090��pfam08708��Rep_trans��pfam02486��Viral_Rep��pfam02407��N��δ������֪Rep���ף�#�����о���������ͬԴ���ƶ���Rep���ס�Note: Rep_1: pfam01446; Rep_3: pfam01051; RepA_C: pfam04796; Rep*: pfam03090 and pfam08708; Rep_trans: pfam02486; Viral_Rep: pfam02407; N: No known Rep has been encoded; #: Putative Rep by sequence similarity in this study.
����2.2�� ˫��˾�����Rep������������
��������Rep���װ��������е�ͬԴ�ԣ�������˫��˾�����Rep���������������ͼ1�����ݽ��������˽ṹ����Ҫ��֧�����Ŷȣ�˫��˾���������Rep�������Ծ���Ϊ7����ͬ�ĵ����壬��������pBIF10�����Rep����(AAZ79481.1)��Rep����������������ǰ��Rep���ױ��ؽṹ�������һ�¡�����Rep�����������Ķ�Ӧ��ϵ��35�����뵥һRep������������Rep��������������ȷ����Ϊ6����ͬ���͵��������壬�����������ͼ���III��˫��˾���Ȼ��������Ҫ���ͺ����Ƽ��壬���ݽ���������III��֧�����˽ṹ����Ҫ��֧�����Ŷȣ�����III�������Խ�һ����ȷ����Ϊ2��������Ⱥ(ͼ1)�����⣬����3�������������ܻ��ֵ���һ���͵��������壬���������ȷ���ֵ��ǵ�һ���͵��������塣���磬��������pDOJH10L����Rep_3 (WP 011067901.1��WP 008783729.1)��RepA_C (WP 080504066.1)����Rep���ף���������ͬʱ��������������III�ͼ���VI (ͼ1)��ʵ���ϣ����������ܿ�������2���������ͬ�������������ں��γɵģ�������������������Σ���ˣ��÷����Ը��������ķ�����ǡ�������˸��������������в�ͬ���������еĽ�����Դ�����һ��֤���÷�����˫��˾������������Ч�Ժ�ȷ�ԡ���֮�������������������˫��˾�����Rep��������������Rep���ױ��ؽṹ��������и��ߵķֱ��ʺ��õķ���Ч�������Զ�˫��˾����б���Rep������Ȼ�������м���Ч��ȷ��ϵͳ���࣬��ˣ�Rep�������п�����Ϊ˫��˾����б���Rep������Ȼ����ϵͳ��������ķ�����ͷ��ӱ�ǡ�
����ͼ1 ˫��˾�������Rep��������
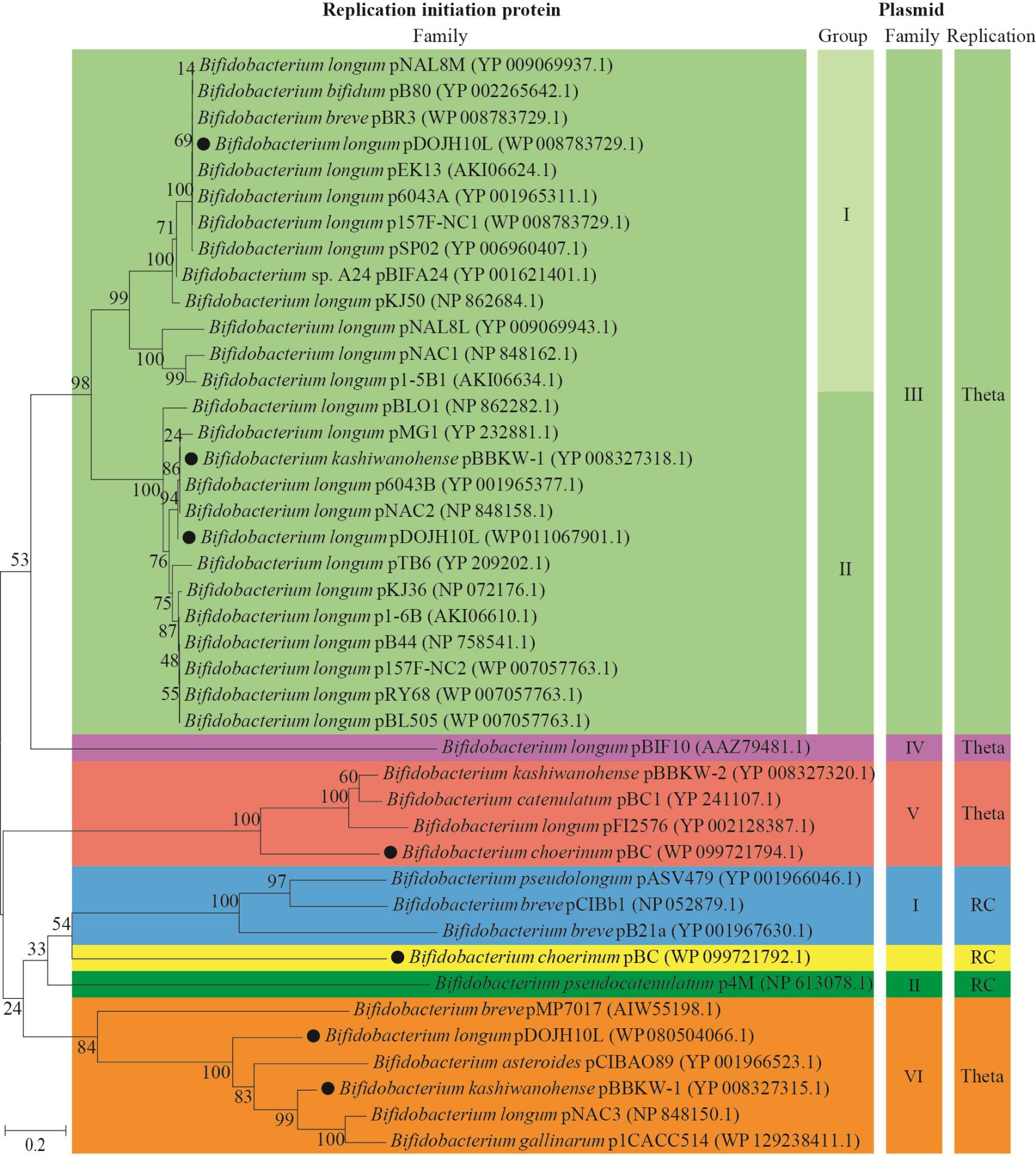
����Figure 1 Phylogenetic tree of the plasmid Reps in Bifidobacterium
����ע����������Rep���ף�RC��Rolling-circle���������ַ�Ϊ��������Rep���װ��������е�¼�ţ���֧�����ֱ�ʾ���Ŷȣ���߱�ʾ�Ŵ����롣Note: Black dot (��) indicates complex plasmid; RC indicates Rolling-Circle; The characters in parentheses are accession numbers of the plasmid-encoded Rep amino acid sequence; Branch point number indicates confidence; Scale indicates evolution distance.
����2.3�� ˫��˾����������鹲���Է���
�������ڸ���������Ȼ����������Ψһ������Ŵ�Ԫ������ˣ����û����鹲���Է��������������ϵͳ����ʱ��ͬʱ������ͬԴ�Ը�������Ϊ��������������Ҫԭ��ͱ����������������鹲���Ժ��������з�����˫��˾���ȫ��41����������ϵͳ��������ͼ2��ʾ��
����ͼ2 ˫��˾������������鹲����
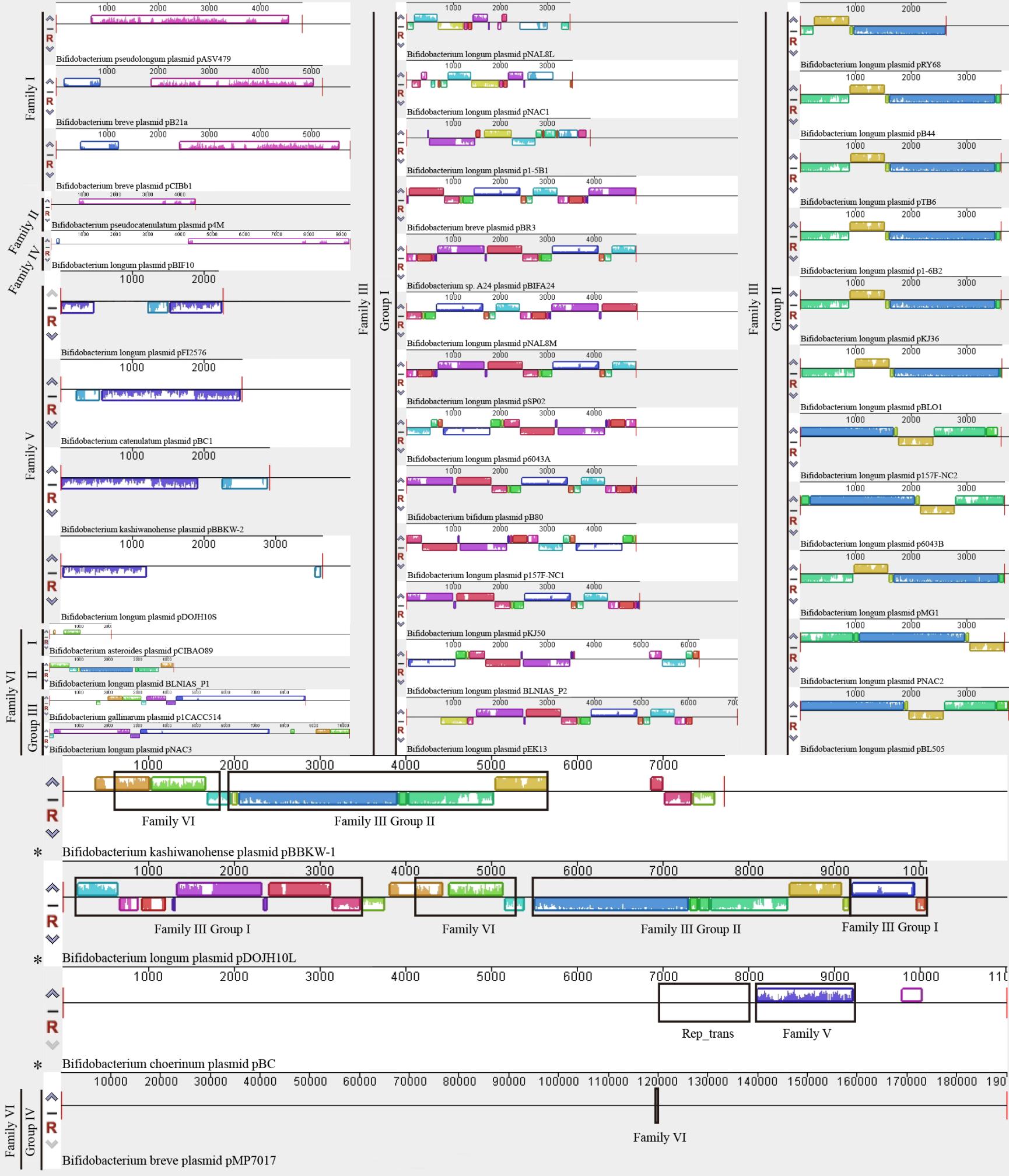
����Figure 2 Genome colinearity of the Bifidobacterium plasmids
����ע��*�������������ڿ�ͬԴ������Note: Asterisk (*) indicates complex plasmid; Black rimmed indicates homology region.
����2.3.1�� ˫��˾�����������I
�����������鹲���Ժ��������з�������������pASV479��pCIBb1��pB21a3���и߶�ͬԴ�ĸ����ӺͱȽϱ��صĻ����鹲���ԣ�������˫��˾�����û����ʾͬԴ������(ͼ2)�����ǵĸ����Ӿ���ͬԴ�ĵ�����㡢˫������rep������ɣ�Rep�������б��ؽṹ��Rep_1 (pfam01446)����Щ��������֪��������pC194��������[22]�����Ӹ߶����ơ�ͬʱ��O’Riordan���о���֤��������pCIBb1���ù������ƽ��и���[23]����ˣ����ڻ����鹲���Ժ�����������������3������������ȷ����Ϊͬһ�������������壬���ù������Ƹ��ƣ�����Ϊ˫��˾�����������I��
����2.3.2 ��˫��˾�����������II
���������鹲���Է�����ʾ������p4Mֻ������pBIF10�ڸ���������������ʾ��һ����ͬԴ�ԣ�������˫��˾�����û����ʾ�κ�ͬԴ��(ͼ2)�����������з�����ʾ������p4M��Rep�����б��ؽṹ��Viral_Rep (pfam02407)����GenBank���ݿ����ڶ����Զ���ֲ�ﵥ��DNA��״������Rep������ʾ�߶�һ���ԡ�Ŀǰ����������p4Mֻ��һƪ���ױ���[24]�����о�����������p4M�����Rep��������ֶ��ﻷ�β��������Rep����ͬ����һ���������Ƶ�������Rep�����壬�ü��嵰�ܿ����ɿ����ֵ������¼����������ڻ����鹲���Ժ�������������������p4M���Ի���Ϊһ���������͵��������壬���ܲ��û��β������ƵĹ������Ƹ��ƣ�����Ϊ˫��˾�����������II��
����2.3.3�� ˫��˾�����������III
�����������鹲���Ժ��������з���������25���������и߶����Ƶĸ����ӣ��������������⣬��Щ���������鹲�������Է�Ϊ2���߶ȱ��ص�������Ⱥ(ͼ2)�����������з�����ʾ����Щ���������Ӿ���rep���������ε��ظ���������ɣ�Rep�������б��ؽṹ��Rep_3 (pfam01051)���ظ��Ӿ���3×10 bp����4×10 bp��(3-5.5)×22 bp���鴮���������ظ�������ɡ���Щ��������֪Theta����pUCL287��������[25]�߶����ơ���ˣ����ڻ����鹲���Ժ�������������Щ����������ȷ����Ϊͬһ�����͵��������壬���ܲ���Theta���Ƹ��ƣ�����Ϊ˫��˾�����������III�����ݻ����鹲���Ե����Բ��죬���Խ�һ������Ϊ2��������Ⱥ�����⣬�����鹲���Է������֣��������ݿ�������BLNIAS_P2û��ע��rep����ʵ���ϸ����������˼���III�����߶�ͬԴ���ظ��Ӻ�rep����ֻ����rep���������������������ͻ�䣬���ܲ���������Rep���ס����⣬���ܼ���III��������������һ�����͵�Theta���������У�������pNAL8L[26]��pNAC1[27]��pKJ36[28]��pMG1[29]��pSP02[30]��pKJ50[28,31]��DNA�ӽ�ʵ������ʾ����Щ�������ƹ����ж����ڵ����м��壬�������м����ǹ������������ĵ�������[32]����ˣ���Щ������ʵ�ĸ������ͺͻ�����Ȼ���ɣ���Щ��������ر���Ҳ���ἰ����������һ��������ʾ������III�������ܴ���һ��δ֪���������ƻ��ơ�
����2.3.4 ��˫��˾�����������IV
���������鹲���Է�����ʾ������pBIF10ֻ�����II����p4M�ڸ���������������ʾ��һ����ͬԴ�ԣ���������˫��˾���������û����ʾͬԴ��(ͼ2)�����������з�����ʾ������pBIF10��������rep���������ε��ظ���������ɣ�Rep�����б��ؽṹ��Rep_3 (pfam01051)���ظ�����3.5×20 bpһ�鴮���������ظ�������ɡ���Щ�����������������pBIF10�ĸ����ӽṹ��Rep���ṹ�������III�������ƣ���������Rep��������(ͼ1)���ظ��ӽṹ�ͻ����鹲����(ͼ2)�������III�������Բ�ͬ���������pBIF10���Ի���Ϊһ���������͵��������壬�Ʋ����Theta���Ƹ���[33]������Ϊ˫��˾�����������IV��
����2.3.5 ��˫��˾�����������V
���������鹲���Ժ��������з�����ʾ��pFI2576��4���������и߶�ͬԴ�ĸ����Ӻ߶ȱ��صĻ���������������˫��˾�����û����ʾͬԴ������(ͼ2)�����ǵĸ�������rep���������εĸ������������ɣ�Rep�������б��ؽṹ��pfam03090��pfam08708����������������֪��Theta����ColE3��������[34]�߶����ơ�ͬʱ������pFI2576[35]��pBBKW-2[8]��pDOJH10S[36]���Ѿ���ʵ��֤ʵ����Theta���ƽ��и��ơ����⣬�����鹲���Է������֣�����pDOJH10Sʵ���ϱ�����rep����ֻ�Ǹû�������ͻ�䣬�γ��˼ٻ��������������鹲���Ժ�������������4������������ȷ����Ϊ�µ��������壬����Theta���Ƹ��ƣ�����Ϊ˫��˾�����������V��
����2.3.6 ��˫��˾�����������VI
���������鹲���Ժ��������з�����ʾ��pCIBAO89��5��������Ȼ�������С�����Բ������ţ������Ǿ���һ��ͬԴ�ĸ���������(ͼ2)���ø�������rep���������ε��ظ���������ɣ�Rep�������б��ؽṹ��RepA_C (pfam04796)���ظ������С��ṹ�ͼ�����ֳ��ϵ͵ı����ԡ�Ŀǰ��5�������ĸ��ƻ���û�еõ�ʵ����֤������������pCIBAO89��pNAC3�ı����Ʋ⣬���ǿ��ܲ���Theta���ƽ��и���[27,37]��������5�������Ļ������С�����Բ������ţ�����������ͬԴ�Ը����ӵĴ��ڱ��������Ǻܿ�������ͬһ������������֮�����˷����������ˣ�����ͬԴ�Ը�������������5���������Ի���Ϊͬһ�����͵��������壬���ܲ���Theta���Ƹ��ƣ�����Ϊ˫��˾�����������VI��ͬʱ�����ڻ����鹲���Ե����Բ��죬���Խ�һ������Ϊ4����ͬ��������Ⱥ(ͼ2)�����⣬�ü��岻������˫��˾�����С������pCIBAO89��Ψһ�ľ�������pMP7017�����Ҹü���������������Ҳ��˫��˾������ģ������������VI�ܿ�����˫��˾���Ȼ�����л���ˮƽת�Ƶ���������¼�������Ƶ�����������塣���⣬������BLNIAS_P2��pDOJH10S���һ���������鹲���Է������֣�����BLNIAS_P1ͬ������һ��rep�ٻ���
����2.3.7�� ˫��˾�����������
���������鹲���Ժ��������з�����ʾ����������pBBKW-1ͬʱ������������III����������VI���͵ĸ����ӣ���������pBC������������VͬԴ�Ը�����֮�⣬��������һ�����ص�Rep_trans����Rep���ף���������pDOJH10L����ͬʱ����3�������ĸ�����(ͼ2)����Щ�����������Щ���������ܿ����ڽ�����2����ͬ��������������������¼��ں��γɣ���������pBBKW-1��pDOJH10L�ı������[8,36]һ�¡�ʵ���ϣ���������ֻ����Ȼ�����������̵�һ������Σ��������泣̬�����ڸ�����������2�����ϲ�ͬ���������ĸ����������ܻ��ֵ���һ���͵��������壬������ǿ�����Ϊ���ص��������ͽ��е������ࡣ
����2.4 ��˫��˾�����Ȼ����ϵͳ������������
�����ۺ�����Rep����ϵͳ�������ͻ����鹲���Է��������˫��˾�������41����Ȼ�������Ի���Ϊ6����ͬ���ͺ���Դ�����������3�����صĸ�������������III�ͼ���VI�����ɽ�һ������Ϊ��ͬ��������Ⱥ(ͼ1��ͼ2)������I��II�������ܲ��ù������Ƹ��ƣ�����III-VI�������ܲ���Theta���Ƹ��ơ�����III������Ա������࣬��˫��˾�����Ȼ��������Ҫ���ͺ����Ƽ��塣����VI������Ա��С�����Ͷ�������ḻ����˫��˾�����Ȼ����ϵͳ������ϵ��ӵ��������塣����������ˮƽ���ַ�����������ȫһ�£����Ǽ���ˮƽ�����鹲���Է��෨���Խ��и���ϸ�µ����ͷ���(�������VI���������ͷ��࣬ͼ2)�����ֶ��������ķ�������Ϊ֤������֤�������Щ���ȷ����Ч�ؽ�ʾ�����з�������֮���ϵͳ������ϵ�Ͷ�Ӧ�ķ���ѧ��λ���ɹ��ؽ�����˫��˾�����Ȼ����ϵͳ���������������ϵ���Խ��˫��˾���Ȼ��������ͼ���������Ҫ�IJο���ֵ��ָ�����塣
����3 ������������
��������ͨ��Rep���������ͻ����鹲�������ַ��������Ŀǰ�Ѳ����˫��˾�����Ȼ����������ϵͳ�����о����о������ʾ������Rep�����������෨�ɽ�˫��˾�����38������Rep������Ȼ��������Ϊ6����ͬ���͵����������3�����صĸ�������������III�����ɽ�һ������Ϊ2��������Ⱥ(ͼ1)�����������鹲���Է��෨�ɽ�˫��˾�����41����Ȼ��������Ϊ6����ͬ���͵����������3�����صĸ�������������III�ͼ���VI�����ɽ�һ������Ϊ2����4��������Ⱥ(ͼ2)������3��rep����ͻ�����������ַ�������������������ˮƽ��ȫһ�£���������֤���˷������ͷ��������Ч�ԡ�ȷ�ԺͿɿ��ԡ��Ƚ����ַ����ķ�������֪�������鹲���Է��෨��ͨ���Ժͷֱ���(���ͷ����ϸ��)��������Rep�����������෨����Щ���֤���������鹲���Է��෨���Զ�˫��˾�������Ȼ��������ȫ�桢ϸ�ºͿɿ���ϵͳ���࣬�����˫��˾���Ȼ����һ�ֱȽ������ϵͳ�������ͬʱ�����������鹲���Է��෨ʹ��ʱ��Ҫͬʱ���������������������������ͱȽϣ���һ���̶��������˷���ʱ��Ͳ����Ѷȣ�������ڳ�ѧ�ߺ��ƻ��Ʋ����������������͡��Ƚ϶��ԣ�����Rep�����������෨���Ӽ����ٺ�ʵ�ã����ʺ������������͵ij�����������֮������Rep���������ͻ����鹲���Է��෨�����ŵ�Ͳ��㣬Ӧ���ݾ�����о�Ŀ�ġ�����ͷ�Χ��������ѡ��һ�ֻ����ַ������Ժͷ�������ﳤ�̣ܶ��ſ��ܻ����ѵ������������������
��������DNA�������Ŀ��ٷ�չ���������������Ѹ�����ӣ������������ͼ�������˸��ߵ�Ҫ��ͬʱҲ��ʹ���ڻ���(��)ϵͳ������ϵ������ϵͳ�����Ϊ���ܡ�ǰ���Ѿ��ᵽ�����ڽ����������������Ա��ͷ��෨�Ѿ�������������ʱ�����������ķ���ͼ����������������������������е�ϵͳ������õ��˽Ͽ�ķ�չ�����и����Ӻ��ɳ�ø���෨����д�����[11,12,13]���Ŵ�ѧ�о���ʾ����������������Ψһ�ձ���ڵ��Ŵ���Ԫ���������������в��������ձ鱣�صĻ��������Ŵ�Ԫ������ɺͽṹ�����Ҷ���[13]����ˣ����ܸ�����������ϵͳ������Ψһ�����ձ��Ժ�ͨ���Ե��Ŵ���ǣ����������������ӷ��෨ֻ��һЩ�к����еõ��˷�չ��Ӧ�ã��������������������о���δ�����������о�ʹ�õ�Rep�����������෨ʵ���������������ӷ��෨�ļ���ʡȥ�˸��������������еķ����ͼ�����ֻ�����˸������������Rep���ף��������˷�����ļ���Ժ�ʵ���ԡ��ɳ�ø������ˮƽת�Ʊ���Ĺؼ�ø��һЩ�о������ɳ�ø��ϵͳ������ϵ�Կ��ƶ�����ʵ���˼���Ч��ϵͳ���࣬���÷������ڲ���ת��������Ч�����Ӱ���˸÷����Ĺ㷺Ӧ��[13]��
������һ���棬�뵥�����ػ������ȣ������ϻ���ȫ����������ͬԴ�Ե�ϵͳ��������и��õ�ȷ�ԺͿɿ��ԣ����Ҿ���������ȫ���Ժ�ͨ���ԡ������о���ʾ�������������Կ�����Ƶ������ˮƽת�ƣ�ͻ��������ʸߣ������ڹ��л������״�������������һֱ�������о�������ѵ����ս��Ŀǰ�������û��һ��ͨ�á�ȷ�ķ�������������˫���������ͽ�Ԫ����ϵͳ����ֱ��������������ࡢ�����ͼ����Ļ��ң������谭������������Ӧ���о��ķ�չ����������Rep���������ͻ����鹲���Է��෨������ǰ�ڶԺ�ھ��ŵ�(G+C)mol%��������˾�����Ȼ���������˳ɹ���ϵͳ����[17,18,19]�����о������ʾ�������ַ��෨ͬ�������ڷ��߾��Ÿ�(G+C)mol%����˫��˾�����Ȼ������ϵͳ���࣬��һ����֤����չ����Щ���������Ч�Ժ�ͨ���ԡ����о���֮ǰ��˾��������о����һ�±��������������鹲���Է��෨��Rep�����������෨���и��ߵ�ȷ�ԺͿɿ��ԣ����ұ��ֳ����õ�ȫ���Ժ�ͨ���ԡ���ˣ��÷����˷������������ӷ��෨(����Rep�����������෨)���ɳ�ø���෨�IJ����ƿ��������������������ϸ����Ȼ������ϵͳ���ࡣ������Ƕȶ��ԣ����������鹲���Է��෨�Ľ����ͷ�չ������Ϊ�����˾���˫��˾���Ȼ������ϵͳ��������ṩ���µķ����������Ϊ����һ��ͨ�á�ȷ��ϸ����Ȼ����������춨����Ҫ���о����������ݡ���ˣ��÷������ܶԽ������ϸ����Ȼ������ϵͳ���ࡢ�����Ԫ�綨��ϵͳ������˫���������ͷ�������о������ձ��ָ�������DZ�ڵ�Ӧ�ü�ֵ��ͬʱ�����������鹲���Է��෨��Ҫ����������Ϣѧ����Mauve�ڵ����н��в�����ʵ���˽ϸߵ�ͨ��(һ���Է�����˫��˾�41����������˾�����5����116��[19])���Զ�����������������˸�ͨ�������д�����������ͼ��������⣬�����˺������ʱ����Ȼ��������ĸ�ͨ�����Զ���������Ȼ�������������鹲���Է��෨�Դ���һЩ���㣬��ʹ�ù����У������ʱ��ͷ���Ч�����д��ڽ�һ���Ż����Ľ�����ߡ�
���������
����[1] Russell DA, Ross RP, Fitzgerald GF, Stanton C. Metabolic activities and probiotic potential of bifidobacteria[J]. International Journal of Food Microbiology, 2011, 149(1): 88-105
����[2] Arboleya S, Watkins C, Stanton C, Ross RP. Gut bifidobacteria populations in human health and aging[J]. Frontiers in Microbiology, 2016, 7: 1204
����[3] Ku S, Park MS, Ji GE, You HJ. Review on Bifidobacterium bifidum BGN4: functionality and nutraceutical applications as a probiotic microorganism[J]. International Journal of Molecular Sciences, 2016, 17(9): 1544
����[4] Fu GF, Li X, Hou YY, Fan YR, Liu WH, Xu GX. Bifidobacterium longum as an oral delivery system of endostatin for gene therapy on solid liver cancer[J]. Cancer Gene Therapy, 2005, 12(2): 133-140
����[5] Tang W, He YF, Zhou SC, Ma YP, Liu GL. A novel Bifidobacterium infantis-mediated TK/GCV suicide gene therapy system exhibits antitumor activity in a rat model of bladder cancer[J]. Journal of Experimental & Clinical Cancer Research, 2009, 28(1): 155
����[6] Ruiz L, Delgado S, Ruas-Madiedo P, Sánchez B, Margolles A. Bifidobacteria and their molecular communication with the immune system[J]. Frontiers in Microbiology, 2017, 8: 2345
����[7] Mahony J, Lugli GA, Van sinderen D, Ventura M. Impact of gut-associated bifidobacteria and their phages on health: two sides of the same coin?[J]. Applied Microbiology and Biotechnology, 2018, 102(5): 2091-2099
����[8] Lee JH, O’Sullivan DJ. Sequence analysis of two cryptic plasmids from Bifidobacterium longum DJO10A and construction of a shuttle cloning vector[J]. Applied and Environmental Microbiology, 2006, 72(1): 527-535
����[9] Orlek A, Stoesser N, Anjum MF, Doumith M, Ellington MJ, Peto T, Crook D, Woodford N, Sarah Walker A, Phan H, et al. Plasmid classification in an era of whole-genome sequencing: application in studies of antibiotic resistance epidemiology[J]. Frontiers in Microbiology, 2017, 8: 182
����[10] Hedges RW, Datta N. Plasmids determining I pili constitute a compatibility complex[J]. Journal of General Microbiology, 1973, 77(1): 19-25
����[11] Carattoli A, Bertini A, Villa L, Falbo V, Hopkins KL, Threlfall EJ. Identification of plasmids by PCR-based replicon typing[J]. Journal of Microbiological Methods, 2005, 63(3): 219-228
����[12] Carloni E, Andreoni F, Omiccioli E, Villa L, Magnani M, Carattoli A. Comparative analysis of the standard PCR-based replicon typing (PBRT) with the commercial PBRT-KIT[J]. Plasmid, 2017, 90: 10-14
����[13] Fernandez-Lopez R, Redondo S, Garcillan-Barcia MP, de la Cruz F. Towards a taxonomy of conjugative plasmids[J]. Current Opinion in Microbiology, 2017, 38: 106-113
����[14] Bertini A, Poirel L, Mugnier PD, Villa L, Nordmann P, Carattoli A. Characterization and PCR-based replicon typing of resistance plasmids in Acinetobacter baumannii[J]. Antimicrobial Agents and Chemotherapy, 2010, 54(10): 4168-4177
����[15] Jensen LB, Garcia-Migura L, Valenzuela AJS, L?hr M, Hasman H, Aarestrup FM. A classification system for plasmids from enterococci and other Gram-positive bacteria[J]. Journal of Microbiological Methods, 2010, 80(1): 25-43
����[16] Lozano C, García-Migura L, Aspiroz C, Zarazaga M, Torres C, Aarestrup FM. Expansion of a plasmid classification system for Gram-positive bacteria and determination of the diversity of plasmids in Staphylococcus aureus strains of human, animal, and food origins[J]. Applied and Environmental Microbiology, 2012, 78(16): 5948-5955
����[17] Sun DQ, Li HF, Song DW, Yang J. Classification of natural plasmids in Lactobacillus plantarum[J]. Food Science, 2017, 38(12): 69-74 (in Chinese)
����[18] Sun DQ, Li HF, Yang J, Song DW. Classification of natural plasmids in Lactobacillus brevis[J]. Food Science, 2018, 39(10): 173-178 (in Chinese)
����[19] Sun DQ, Song DW, Li HF. Research on the Genomics of Natural Plasmids of Lactobacillus[M]. Beijing: China Light Industry Press, 2018: 187-189 (in Chinese)
����[20] Kumar S, Stecher G, Tamura K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets[J]. Molecular Biology and Evolution, 2016, 33(7): 1870-1874
����[21] Darling ACE, Mau B, Blattner FR, Perna NT. Mauve: multiple alignment of conserved genomic sequence with rearrangements[J]. Genome Research, 2004, 14(7): 1394-1403
����[22] Seery LT, Nolan NC, Sharp PM, Devine KM. Comparative analysis of the pC194 group of rolling circle plasmids[J]. Plasmid, 1993, 30(3): 185-196
����[23] O'Riordan K, Fitzgerald GF. Molecular characterisation of a 5.75-kb cryptic plasmid from Bifidobacterium breve NCFB 2258 and determination of mode of replication[J]. FEMS Microbiology Letters, 1999, 174(2): 285-294
����[24] Gibbs MJ, Smeianov VV, Steele JL, Upcroft P, Efimov BA. Two families of Rep-like genes that probably originated by interspecies recombination are represented in viral, plasmid, bacterial, and parasitic protozoan genomes[J]. Molecular Biology and Evolution, 2006, 23(6): 1097-1100
����[25] Benachour A, Frère J, Novel G. pUCL287 plasmid from Tetragenococcus halophila (Pediococcus halophilus) ATCC 33315 represents a new theta-type replicon family of lactic acid bacteria[J]. FEMS Microbiology Letters, 1995, 128(2): 167-175
����[26] Guglielmetti S, Karp M, Mora D, Tamagnini I, Parini C. Molecular characterization of Bifidobacterium longum biovar longum NAL8 plasmids and construction of a novel replicon screening system[J]. Applied Microbiology and Biotechnology, 2007, 74(5): 1053-1061
����[27] Corneau N, ?mond ?, LaPointe G. Molecular characterization of three plasmids from Bifidobacterium longum[J]. Plasmid, 2004, 51(2): 87-100
����[28] Park MS, Lee KH, Ji GE. Isolation and characterization of two plasmids from Bifidobacterium longum[J]. Letters in Applied Microbiology, 1997, 25(1): 5-7
����[29] Park MS, Moon HW, Ji GE. Molecular characterization of plasmid from Bifidobacterium longum[J]. Journal of Microbiology and Biotechnology, 2003, 13(3): 457-462
����[30] ?lvarez-Martín P, ?ycka-Krzesi��ska J, Bardowski J, Mayo B. Sequence analysis of plasmid pSP02 from Bifidobacterium longum M62 and construction of pSP02-derived cloning vectors[J]. Plasmid, 2013, 69(2): 119-126
����[31] Park MS, Shin DW, Lee KH, Ji GE. Sequence analysis of plasmid pKJ50 from Bifidobacterium longum[J]. Microbiology, 1999, 145(3): 585-592
����[32] Ruiz-Masó JA, Machón C, Bordanaba-Ruiseco L, Espinosa M, Coll M, del Solar G. Plasmid rolling-circle replication[J]. Microbiology Spectrum, 2015, 3(1): PLAS-0035-2014
����[33] Ma YP, Xie TT, Hu QW, Qiu ZY, Song FZ. Sequencing analysis and characterization of the plasmid pBIF10 isolated from Bifidobacterium longum[J]. Canadian Journal of Microbiology, 2015, 61(2): 124-130
����[34] Aoki K, Shinohara M, Itoh T. Distinct functions of the two specificity determinants in replication initiation of plasmids ColE2-P9 and ColE3-CA38[J]. Journal of Bacteriology, 2007, 189(6): 2392-2400
����[35] Moon GS, Wegmann U, Gunning AP, Gasson MJ, Narbad A. Isolation and characterization of a theta-type cryptic plasmid from Bifidobacterium longum FI10564[J]. Journal of Microbiology and Biotechnology, 2009, 19(4): 403-408
����[36] Takahata M, Toh H, Nakano A, Takagi M, Murakami M, Ishii Y, Takizawa T, Tanabe S, Morita H. Complete sequence analysis of two cryptic plasmids from Bifidobacterium kashiwanohense JCM 15439 (type strain) isolated from healthy infant feces[J]. Animal Science Journal, 2014, 85(2): 158-163
����[37] Cronin M, Knobel M, O'Connell-Motherway M, Fitzgerald GF, van Sinderen D. Molecular dissection of a bifidobacterial replicon[J]. Applied and Environmental Microbiology, 2007, 73(24): 7858-7866





