社会交往是个体在社会群体里必不可少的沟通方式,有助于个体成长和后代繁衍。即使简单的社会交往行为也需要神经系统精确的分析、整合外界信息并调节运动系统使行为正常进行。许多精神-神经性疾病 ( 如孤独症、抑郁症等) 的患者都表现出不同程度的社会交往障碍。啮齿类动物的社会交往主要依靠嗅觉系统来进行信息交流,其嗅觉系统分为主嗅觉系统和犁鼻系统。社会交往行为中,主嗅觉系统主要探测空气中传播的挥发性化学物质,而梨鼻系统探测和处理动物释放的信息素引起动物产生相应的行为反应。采用化学方法或破坏嗅上皮或者手术切除犁鼻器官导致嗅觉系统功能不完整时,动物的异性别社会交往行为表现异常。同性别社会交往在社会交往行为中占有重要比例,但是目前关于啮齿类动物的同性别社会交往行为研究较少。究竟有哪些脑区可能参与了此类交往活动鲜有报道。细胞外信号调节激酶 ( extracellular signal-reg-ulated kinase 1 /2,ERK1 /2) 作为调节转录因子活性的重要信号分子,通过磷酸化快速将细胞外信号转至细胞内调节基因转录。pERK1/2 可被作为神经元迅速激活的一个可靠的标志物研究与动物某行为有关的脑区。本实验用三箱室社会交往箱观察雄性大鼠同性别社会交往行为,同时运用免疫组织化学观察 pERK1/2 在 大 鼠 脑 内 表 达 的 变 化 以 探 讨pERK1 /2 在大鼠同性别社会交往中的作用。
材料和方法
1 材料
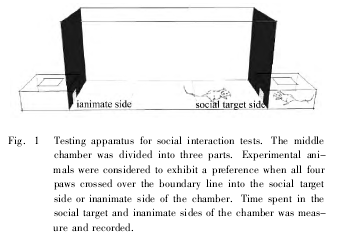
健康清洁级成年雄性 SD 大鼠,体重220 ~260 g,由中南大学实验动物学部提供。实验前所有大鼠均饲养在动物室适应环境一周后开始实验。社会交往箱如图 ( Fig. 1) 所示。将雄性大鼠分组进行正常情况下同性别社会交往实验和嗅觉剥夺后同性别社会交往实验。两组实验分别设置对照组: 社会交往箱中两侧无交往对象鼠时在箱中活动情况 ( 非社会交往组) 和用生理盐水滴鼻后进行社会交往实验。
2 方法
2. 1 同性别社会交往行为检测方法 在社会交往行为检测前,待测雄鼠于每天下午 2∶ 00 置于社会交往箱中间的大有机玻璃箱内 8 min 适应环境,每日 1 次,连续 4 d。社会交往行为检测时,先将正常雄性大鼠随机的放在社会交往箱的一侧小箱子里,另一端的小箱子保持空白,再将待测鼠放在中间大箱子的正中间的区域,摄像机记录 10 min 内待测鼠在社会交往箱社会交往侧 ( 含有正常大鼠一侧) 和无生命体侧 ( 空白箱一侧) 停留的时间、接触两端金属网的次数。当雄鼠四足都跨出社会交往箱底部的中间区域开始记录雄鼠在社会交往箱左、右两个区域停留的时间,当雄鼠任意一足跨入社会交往箱底部正中间区域时停止记时。每只雄鼠检测结束后,将其放回原笼,清扫粪便,并用70% 酒精去除社会交往箱内的残余气味。
2. 2 硫酸锌滴鼻嗅觉剥夺方法 本 实验采用10% 硫酸锌破坏雄鼠的嗅上皮,研究主嗅觉系统在雄鼠同性别社会交往行为中的作用。用 10% 水合氯醛将动物麻醉后,仰卧位置于手术台上,两侧鼻孔内滴注 10% 硫酸锌溶液 ( 天津市博迪化工股份有限公司) 直至有液体从鼻孔内流出为止,对照组雄鼠两侧鼻孔内滴入生理盐水。待动物完全苏醒后放回动物室。动物嗅觉剥夺后第 1 d 进行嗅觉检测,确定嗅觉功能丧失后进行社会交往实验。
2. 3 免疫组织化学染色 行为实验结束后立即10% 水合氯醛麻醉,用生理盐水灌注后再用 4℃ 的4% 多聚甲醛固定液固定,取脑于 4°冰箱后固定过夜,15%、30% 蔗糖脱水后做连续冠状面冰冻切片,切片厚度 30 um。切片用 3% 过氧化氢溶液处理 16 min 以阻断内源性过氧化物酶的影响,0. 1%Triton X-100 的 5% BSA 室 温 封 闭 2 h, 一 抗pERK1 /2 ( 兔抗,1 ∶ 1000,Cell Signaling) 室温孵育 2 h 后,4℃冰箱中孵育过夜。第 2 天,复温 30min 后,加生物素化羊抗兔 IgG ( 1 ∶ 200,Vector) ,室温孵育 2 h 后,过氧化物酶标记的 ABC 复合物( A 液和 B 液/1∶ 200,提前 30 min 配制) 室温孵育2 h 后,DAB 液显色,第 2 天开始每步骤之前用 0.01 mol / L PBS 振荡漂洗 8 min × 3 次。
统计学处理: 采用 Nikon H600L 显微镜下观察pERK1 /2 免疫阳性细胞表达的脑区。采集 10 倍图像,HPIAS-1000 高清晰度彩色病理图像分析系统( 同济医科大学千屏影像工程公司) 统计每平方毫米免疫阳性细胞数。所得数据均以 Mean ± SD 表示,采用 SPSS 13. 0 软件和 Graphad prism5. 0 进行统计分析,Levene 检验进行方差齐性检验 ( 方差齐性检验水准为 0. 10) ,Two-Way ANOVA 进行两样本均数的比较,多个样本均数的比较采用单因素方差分析 ( One-Way ANOVA) ,多个样本均数间的多重比较采用 post hoc Dunnett 检验。P 值小于 0.05 认为差异具有统计学意义。
结 果
1 正常雄性大鼠同性别社会交往行为
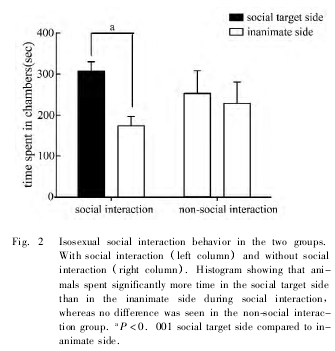
当社会交往箱左右两端都没有放入交往对象鼠时,正常雄鼠在社会交往箱两侧停留的时间差异无统计学意义 ( t = 0. 4136,P > 0. 05) 。当一侧有交往对象鼠时,大鼠在社会交往侧停留的时间明显多于在无生命体侧停留的时间 ( t = 3. 639,P < 0.001,Fig. 2) 。
2 正常雄鼠与同性别大鼠社会交往后脑内 pERK1 /2表达
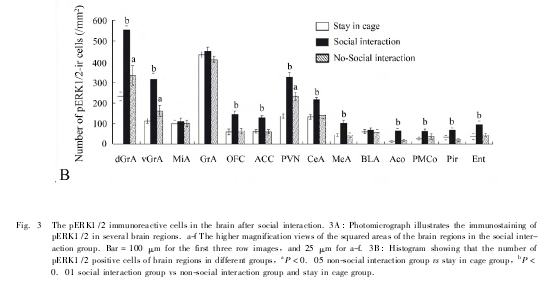
正常雄鼠与同性别大鼠社会交往后,多个脑区内存在 pERK1/2 的阳性表达: 与空白对照组和非社会交往组雄鼠相比,社会交往组雄鼠在与主嗅觉系统相关的脑区 ( 如主嗅球 dGrA、vGrA,梨状皮质 Pir、内嗅皮质 Ent) 以及室旁核 PVN、眶额皮质OFC、前扣带皮质 ACC、杏仁中央核 CeA 与杏仁内侧核 MeA) 内 pERK1/2 阳性细胞表达数量明显增加 ( P <0. 01) ,在副嗅球僧帽细胞层 MIA 和颗粒层 GrA 内 pERK1/2 阳性细胞表达数量差异无显着性 ( P >0. 05) 。
3 ZnSO4 嗅觉剥夺后社会交往行为及脑内 pERK1 /2表达变化
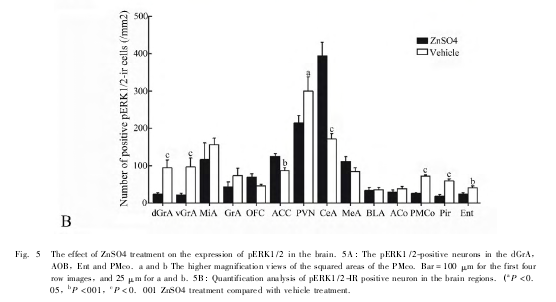
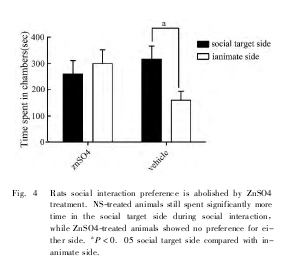
鼻腔滴生理盐水的对照组大鼠在社会交往测停留的时间明显多于无生命体侧 ( P <0. 05) ,而嗅觉被剥夺后大鼠在社会交往箱中社会交往侧和无生命体侧停留的时间差异无统计学意义 ( P > 0. 05,Fig. 4) 。雄鼠脑内 pERK1 /2 在主嗅觉系统相关脑区 ( 主嗅球、内嗅皮质、梨状皮质) 表达明显下降,在杏仁中央表达明显增加 ( Fig. 5) 。
谈论
Crawley 等人进行小鼠的社会交往实验时发现小鼠存在明显的社会交往倾向,但是未探究参与社会交往行为的神经调节机制。本实验采用三箱室社会交往箱研究大鼠同性别社会交往行为,以pERK 作为标志物研究参与社会交往相关脑区。
同样地,我们发现当社会交往箱两侧都空箱,大鼠放于社会交往箱中时,大鼠未表现出左右两侧的位置选择偏好。当一侧小箱放有交往对象鼠时,大鼠在社会交往箱中表现出明显的社会交往偏好。10min 社会交往行为后大鼠脑内与主嗅觉系统相关的脑区 ( 如主嗅球、梨状皮质、内嗅皮质) 、室旁核、杏仁中央与内侧核 pERK1/2 表达明显上调,而副嗅球 pERK1/2 的表达在社会交往组与非社会交往组差异无显着性。Dudley 等人发现主嗅球、副嗅球、杏仁核及前扣带皮质在啮齿类的异性别社会交往中被激活。这提示梨鼻系统在异性别社会交往中的发挥特异性作用,而主嗅觉系统参与在同性别社会交往行为作用。硫酸锌可以破坏雄鼠的嗅粘膜,引起动物主嗅觉系统功能暂时性丧失。
但是不影响动物梨鼻系统功能和情绪反应。我们用硫酸锌滴鼻破坏大鼠的主嗅觉系统,嗅觉剥夺后雄鼠在社会交往箱中社会交往偏好现象消失,表明主嗅觉系统被破坏将影响大鼠的同性别社会交往行为,同时大鼠与主嗅觉系统相关的脑区 pERK1/2表达下降。
有多个文献报道 ERK1/2 参与动物社会交往行为: SL327 ( MEK 抑制剂抑制 ERK 磷酸化) 处理的小鼠表现为社会交往行为减少,ERK2 基因敲除后小鼠也表现为社会交往行为减少,转基因雌性小鼠前额叶皮质过表达 ERK2,社会交往行为增加。表明 pERK1/2 参与了小鼠社会交往行为,本实验同性别社会交往行为后主嗅觉相关脑区pERK1 /2 表达上调亦提示 pERK1 /2 可能参与大鼠同性别社会交往行为。MAPK 信号通路在突触形成和个体学习记忆中具有重要作用。有研究表明嗅觉记忆形成期间,嗅球 MEK1 表达上调。主嗅觉系统信号的转导有赖于 III 型 cAMP 酶和主要控制Ca 离子进出的环核苷酸门控通道 ( CNP)。在大鼠社会交往时,信号通过主嗅觉系统细胞内cAMP 转导和 CNP 激活, 参与调节社会偏好行为。海马在记忆形成中,cAMP 和 CNP 信号转导激活下游 ERK 信号通路。主嗅觉系统细胞内cAMP 转导和 CNP 可能激活下游 MAPK 信号通路对个体识别和记忆并调节大鼠交往行为。然而主嗅觉系统 pERK1/2 在大鼠同性别社会交往中的作用需要进一步的实验进行证实。
本实验用 pERK1/2 为标志物检测大鼠同性别社会交往行为后各脑区激活状况,发现与主嗅觉系统相关的脑区被激活。应用 ZnSO4 破坏大鼠主嗅觉系统导致大鼠同性别社会交往行为降低,主嗅觉系统相关脑区激活程度下降,提示主嗅觉系统相关脑区与大鼠同性别社会交往行为存在密切联系。
参 考 文 献
[1] Bateman C. Autism-mitigating a global epidemic [J]. S Afr MedJ,2013,103: 276 - 277.
[2]徐 杨,曹文宇,李昌琪,等. 嗅觉系统在啮齿类动物社会交往行为中的作用 [J]. 中华行为医学与脑科学杂志,2012,21: 956 - 957.
[3]Yokosuka M. Histological properties of the glomerular layer in themouse accessory olfactory bulb [J]. Exp Anim,2012,61: 13 -24.
[4]Keller M,Douhard Q,Baum M J,et al. Destruction of the main ol-factory epithelium reduces female sexual behavior and olfactory in-vestigation in female mice [J]. Chem Senses,2006,31: 315 -323.
[5]Feierstein C E. Linking adult olfactory neurogenesis to social behav-ior [J]. Front Neurosci,2012,6: 173.
[6]Taziaux M,Keller M,Balthazart J,et al. Rapid activation of phos-phorylated mitogen-activated protein kinase after sexual stimulationin male mice [J]. Neuroreport,2011,22: 294 - 298.
[7]Niu H,Zheng Y,Huma T,et al. Lesion of olfactory epithelium at-tenuates expression of morphine-induced behavioral sensitization andreinstatement of drug-primed conditioned place preference in mice[J]. Pharmacol Biochem Behav. 2013,103: 526 -534.
[8]Yang M,Crawley J N. Simple behavioral assessment of mouse olfac-tion [J]. Curr Protoc Neurosci,2009,Chapter 8: 8 - 24.
[9]Kaidanovich-Beilin O,Lipina T,Vukobradovic I,et al. Assessmentof social interaction behaviors [J]. J Vis Exp. 2011 ( 48) .
[10]Roullet F I,Crawley J N. Mouse models of autism: testing hypothe-ses about molecular mechanisms [J]. Curr Top Behav Neurosci,2011,7: 187 - 212.
[11]Dudley C A,Chakravarty S,Barnea A. Female odors lead to rapidactivation of mitogen-activated protein kinase ( MAPK) in neuronsof the vomeronasal system [J]. Brain Res,2001,915: 32 - 46.
[12]Martinez-Ricos J,Agustin-Pavon C,Lanuza E,et al. Role of thevomeronasal system in intersexual attraction in female mice [J].Neuroscience,2008,153: 383 - 395.
[13]Satoh Y,Endo S,Nakata T,et al. ERK2 contributes to the controlof social behaviors in mice [J]. J Neurosci,2011,31: 11953 -11967.
[14]Valjent E,Corvol J C,Trzaskos J M,et al. Role of the ERK path-way in psychostimulant-induced locomotor sensitization [J]. BMCNeurosci,2006,7: 20.
[15]郑君芳,华 琳,张秋霞,等. 嗅觉记忆相关蛋白的蛋白质组学研究 [J]. 高等学校化学学报,2010,21: 1535 -1540.
[16]Zibman S,Shpak G,Wagner S. Distinct intrinsic membrane prop-erties determine differential information processing between mainand accessory olfactory bulb mitral cells [J]. Neuroscience,2011,189: 51 - 67.
[17]Ma M. Odor and pheromone sensing via chemoreceptors [J]. AdvExp Med Biol,2012,739: 93 - 106.
[18]Spehr M,Kelliher K R,Li X H,et al. Essential role of the mainolfactory system in social recognition of major histocompatibilitycomplex peptide ligands [J]. J Neurosci,2006,26: 1961 -1970.
[19] Xia Z,Storm D R. Role of signal transduction crosstalk betweenadenylyl cyclase and MAP kinase in hippocampus-dependent memo-ry [J]. Learn Mem,2012,19: 369 - 374