
蹄盖蕨科( Athyriaceae) 为中小型、少数为大型土生蕨类植物,约有 20 属 500 种,广布全世界热带至寒温带各地,尤以热带与亚热带山地为多。我国各属均产,约 400 种。蕨类植物孢子萌发和配子体发育的特性可以为分类学和种的研究提供有效的依据。近年来,对蹄盖蕨科配子体发育过程已有一些研究,在此基础上可知蹄盖蕨科配子体发育的一些特点: 孢子通常大多数种类有周壁,有周壁的孢子外壁通常光滑,无周壁的孢子外壁有颗粒状的、微皱的或刺状纹饰( 很少是光滑的) ,周壁是光滑的到有颗粒的或者少数是刺状的,通常形成短的脊状褶皱; 孢子萌发类型为向心型; 原叶体发育类型是铁线蕨型或近槲蕨型; 蹄盖蕨属( Athyrium) 和双盖蕨属( Diplazium) 的成熟原叶体裸露或有毛状体,冷蕨属( Cystopteris) 具毛状体; 毛状体只在原叶体发育晚期才形成,通常是乳突状分泌型毛状体; 双盖蕨属的一些种类精子器的开裂是由盖细胞形成一个孔状的开口,其它的种类盖细胞破碎,精子器裂开。对于蹄盖蕨科这样一个大科来说,有关配子体发育的研究还是相对薄弱的。
本文对蹄盖蕨科的尖头蹄盖蕨 ( Athyriumvidalii) 、长柄蹄盖蕨( A. longius) 、喜马拉雅蹄盖蕨( A. fimbriatum) 和亮毛蕨( Acystopteris japonica) 配子体的形态和发育过程进行观察,详细描述其配子体在各发育阶段的形态结构特征,为蹄盖蕨科植物的分类和演化研究提供配子体方面的资料。
1、 材料与方法
1. 1 孢子的收集
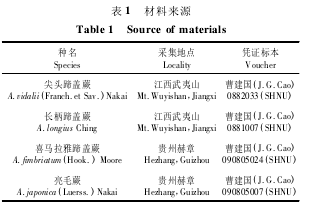
实验用孢子的采集信息详见表 1。采集具有成熟孢子且孢子囊未开裂的叶片,将叶片自植物体上采下后放入信封中,待孢子囊自然裂开,成熟孢子散落于信封中,置 4℃冰箱待用。
1. 2 孢子的接种培养
将成熟孢子用 5% 次氯酸钠溶液灭菌,无菌水清洗后制成孢子悬浮液,并均匀接种在改良 knop’s培养基上。将接种后的培养皿置于 RXZ 型智能人工气候箱中培养。培养条件为全天光照,光照强度约 25 μmol·m- 2·s- 1; 温度约 25℃; 相对湿度约 60% ~ 70%。定期观察,用 Motic BA310 型光学显微镜照相记录。每次培养 6 个小培养皿,共培养 3 次。
2、 结果
通过对尖头蹄盖蕨、长柄蹄盖蕨、喜马拉雅蹄盖蕨和亮毛蕨配子体的形态和发育过程进行观察,将 4 种蹄盖蕨科植物配子体各发育阶段的主要特征总结于表 2。
4 种蹄盖蕨科植物的孢子都为单裂缝,两侧对称,黄褐色; 孢子大小从 26 ×35 μm( 长柄蹄盖蕨)到 36. 4 ×50 μm( 喜马拉雅蹄盖蕨) 不等,孢子表面纹饰为网状、拟网状、脊状褶皱和棒状。接种后6 ~ 14 d 孢子开始萌发,萌发的标志都是首先长出一条初生假根,孢子萌发类型都为向心型。由原叶体原始细胞进行横向分裂开始,经过一系列的横分裂最终形成单列丝状体,丝状体有 2 ~6 个细胞的长度,细胞圆桶状或长圆桶状。此时,丝状体的前端细胞斜向分裂产生顶端细胞。丝状体基部都有孢壁附着。
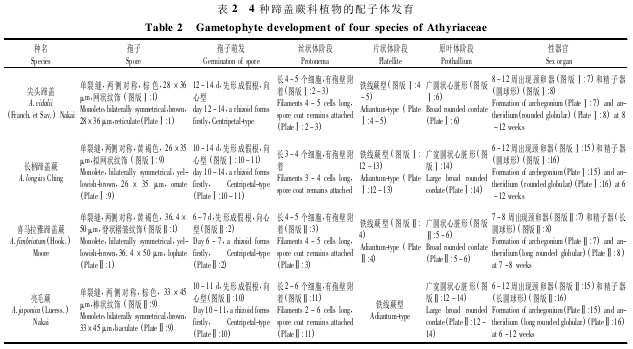
顶端细胞向两侧分裂产生片状体,片状体多为带状。随着原叶体的生长,顶端细胞被分生组织代替,分生组织位于片状体前端中央,分生组织细胞左右对称地分裂形成幼原叶体。幼原叶体为对称的长心脏形,基部楔形,边缘近全缘,分生组织区域也由最初的平直状移至心形凹陷的底部。
接种 6 ~12 周后,在分生组织持续的分裂下,最终发育为成熟原叶体。成熟原叶体为对称的广圆状心脏形至广宽圆状心脏形,宽度等于或大于长度,分生组织两侧边缘多重叠,原叶体边缘全缘或略有波折。原叶体的发育类型为铁线蕨型。成熟原叶体上着生着颈卵器和精子器,颈卵器和精子器都属于薄囊蕨型。颈卵器位于生长点下方至原叶体中部的中脉范围内,颈部有 4 列细胞,每列由 4 ~ 5 层细胞构成,朝向配子体基部略弯曲。精子器产生于原叶体基部的表面和边缘,圆球形或长圆球形,直径 24 ~ 27 μm( 亮毛蕨) 到55 ~ 60 μm( 尖头蹄盖蕨) ,由基细胞、环细胞和盖细胞组成,其中环细胞最大,占精子器的 2/3 ~ 3/4,其次是基细胞和盖细胞。
4 种植物的假根幼时无色,偶有叶绿体; 老时无叶绿体,其顶端常呈棕褐色,有顶端膨胀的现象( 图版Ⅰ: 5,18) ,长柄蹄盖蕨和亮毛蕨假根偶有螺旋弯曲( 图版Ⅰ: 18,图版Ⅱ: 17) 。
3、 讨论
本文观察蹄盖蕨属的尖头蹄盖蕨、长柄蹄盖蕨、喜马拉雅蹄盖蕨与中华蹄盖蕨 ( Athyriumsinense) 、带岭蹄盖蕨( A. dailingense) 、多齿蹄盖蕨( A. multidentatum) 和日本蹄盖蕨 ( A. niponi-cum)的孢子都为单裂缝,两侧对称,孢子萌发类型为向心型,配子体的发育类型为铁线蕨型,精子器和颈卵器都为薄囊蕨型,成熟原叶体为对称的心脏形,无毛状体,这些共同特点可以作为蹄盖蕨属配子体的发育特征,与 Nayar所给出的描述是一致的。但这些种之间在孢子纹饰、孢子萌发和配子体发育速度、丝状体长度、片状体和原叶体的形态以及性器官大小等方面存在明显差异; 且与 Nayar认为的“蹄盖蕨属中一些种类有毛状体”不一致,这可能是种间差异所致。长柄蹄盖蕨、喜马拉雅蹄盖蕨和亮毛蕨的孢子大小、对称性和纹饰类型等孢子形态特征与王全喜等和常缨等的观察相一致。
一些学者认为亮毛蕨属与冷蕨属关系密切,应为后者的一个亚属。将本文观察的亮毛蕨与林孝辉等观察的冷蕨属的山冷蕨( Cystopteris sudet-ica)以及 Nayar的观察比较可知,两属虽然有些性状比较相似,如孢子萌发类型、丝状体形态和性器官类型等,但仍然存在较大差异,如原叶体发育类型、毛状体的有无、成熟原叶体的形态和性器官的大小等,故认为从配子体的发育来看,仍然应该将亮毛蕨独立为一属,支持秦仁昌系统。
王玛丽等依据分子系统学的研究结果,再结合孢子囊群的特征,将蹄盖蕨科划分为 5 个亚科,即冷蕨亚科( Cystopterioideae) 、蹄盖蕨亚科( Athy-rioideae) 、对囊蕨亚科( Doparioideae) 、双盖蕨亚科( Diplazioideae ) 和轴果蕨亚科 ( Rhachido-soroideae) ; 其中冷蕨亚科包括亮毛蕨属 ( Acystop-teris) 、冷蕨属( Cystopteris) 、光叶蕨属( Cystoathyri-um) 和羽节蕨属( Gymnocarpium) 4 个属; 蹄盖蕨亚科包括蹄盖蕨属( Athyrium) 、角蕨属( Cornopteris) 、假冷蕨属( Pseudocystopteris) 、拟鳞毛蕨属( Kuni-watsukia) 及安蕨属( Anisocampium) 5 个属。亮毛蕨的原叶体发育为无毛状体的铁线蕨型与同亚科具毛状体的三叉蕨型的冷蕨属和羽节蕨属截然不同,而与蹄盖蕨亚科的蹄盖蕨属、角蕨属和假冷蕨属是一致的。因此,从配子体世代特征看,亮毛蕨属归于蹄盖蕨亚科较为合适。
参考文献:
1. 王中仁,张宪春,朱维明,等. 中国植物志第三卷第二分册[M]. 北京: 科学出版社,1990.





