
����0������
����������ѧϰ������������ȷ�������Ҫ���ã���������ϵͳ�����Բ���������ء�����������������������ͨ��ϸ����ֳ�������Э�����õ��ڵġ�DCX�������ĺ��ʱ������ݱ�������ֳ��ǰ��ϸ����������ǿɷ�ӳ����ˮƽ�ߵ͡�Caspase-3��ϸ����������Ҫִ���ߣ�Ҳ���ж�ϸ��������������־(biochemi-calmarker)��Ŀǰ��Ӧ�û����ó���ת������ģ���о��������Լ�������C57/BL6С��Ϊ�������壬���й��亣������������ϸ����ֳ�������ϵͳ�о����ʼ���������ʵ��Ӧ��������֯��ѧȾɫ������ӫ��������ӡ�Ǽ��������羵������C57/BL6С��������������ϸ����ֳ������ı仯���ɽ���ϵͳ�Ͷ�̬�۲��о���Ϊ����̽�ֺ����������������Լ������������Լ����Ļ����о��ṩ������ʵ�����ݡ�
����1�������뷽��
����1.1ʵ�鶯�������뺣���걾�Ʊ�
�������꽡��C57/BL6С�����۱���1��2ͬ���������Դ�������˨���ּ�Ϊ����(E)0d;���������������ʱ���Ϊ����(P)0d��ѡȡ����12��14��16��18d̥�������1��3��5��7��14��21��28d��2��3m������ÿ��8ֻ������4%��ͱ����Ƹ�ǻ�������ʸ�ȡ��̥��E10��12��14d��̥��ȡȫ�ߣ������������̥��������ͷȡ�ԣ�4%��ۼ�ȩ����̶�����ˮϴ�����ϵ���Ҵ���ˮ�����ױ�����ʯ�����������5μm��������Ƭ�������黯������ӫ��Ⱦɫ���á�����ȡ�������ʺ�����֯���ڣ�80�汣�棬��������ӡ������������ȡE12��16d��P1��P7d������֯�г�Լ1mm3С�飬��3%���ȩ���½���̶�48h���ھ�1%����̶�1h������羵�걾�Ʊ���Epon812������֬�������LKB-V�ͳ�����Ƭ���뱡��Ƭ�����1μm���ױ�����Ⱦɫ���⾵�۲캣��������λ�����г�����Ƭ����70nm���ؽ���˫Ⱦɫ��EX1200��羵�۲졣
����1.2DCX������֯��ѧȾɫ
��������ʯ����Ƭ�����黯Envision������Ƭ����������ˮ��3%����������Һ����30min��ȥ����Դ�Թ�������ø����ѹ����ԭ����һ��(����DCX1��200��SantaCruz��˾)4���ҹ���μ������ر�ǵĶ���(�ÿ���IgG��1��200��������ɼ������������˾)���·���10min���μ���ù����������-��������ø��Һ���·���10min��DAB��ɫ����ľ�غ˸�Ⱦ����ˮ����������������Ƭ���⾵�¹۲졣���Զ�����PBS����һ����DCX���Կ������غ�ɫ��λ��Ǩ���е���Ԫ�����Լ��ֻ��е���Ԫ��ͻ��
����1.3Caspase-3����ӫ��Ⱦɫ
����ʯ����Ƭ����ˮ��������10min;�ӷ����߶���Ѫ�����·���10min;��һ��(�ÿ���Caspase-3��1��200��SantaCruz��˾)4���ҹ;��FITC���ɽ����IgG(������ɼ������������˾)37��ܹ����30min��65%���ͷ�Ƭ��ӫ�������¹۲죬Caspase-3���Կ�����������ɫӫ�⣬�ֲ���ϸ���������Զ�����PBS����һ����
����1.4Westernblotting���DCX��Caspase-3���ױ���
���������䶳������֯����ϸ���ѽ�Һ��0���¼���ͳ���������20s����30min��4����12000r/min����20min����ȡ����Һ��BCA���ⶨ������������ÿ20μL��50μg�����Ƴ���Ʒ����20����䱣�档ȡ���Ʊ���Ʒ���������Ӿ�ۣ�100V��ѹ�µ�Ӿ��תĤ�������°��תӡ��TBST��ϴ��5%СţѪ��������·��1��2h���ֱ����һ��(DCX��caspase-3���壬ϡ�Ͷ�1��200)4�������ҹ��TBST����Һϴ��3�Σ�ÿ��5min;�ֱ����������·���1h��TBST����Һϴ��3�Σ�ÿ��5min��ECL������ɫ;����β-actin��Ϊ�ڲΣ�ɨ���Ӿ������������������������������Ŀ����������ȡ�����Ŀ���������ڲ���������ȵı�ֵ��ʾ���⵰������
����1.5ͳ�Ʒ���
����ʵ��������ͳ������SPSS16.0����������������x±s��ʾ��������������ݽ����ظ��������ϵķ��������P��0.05Ϊ������ͳ��ѧ���塣
����2�����
����2.1DCX�����黯Ⱦɫ���
����DCX�������Է�Ӧ������غ�ɫ��E12d����ϸ�������ں����ṹԭ���У�E16��E18dCA������ɼ���ɢ�ֲ�������ϸ����P1d��P14d����ϸ������������ų�һ���ܴ���P1d�����P14d��ʧ;���Ӳ㼰���β��е�����ϸ����P1��P7d���٣�P14d����ʧ��P1��P7d����ϸ����ɢ�ֲ��ڳ�״�ظ��㣬P14d��Ҫ�ֲ��ڿ�������1/2��P21d��Ҫ�ֲ����ǿ���ϸ���㣬P28d��3m�ǿ���ϸ��������ϸ������(ͼ1)��

����2.2Caspase-3����ӫ��Ⱦɫ���
����Caspase-3���Է�Ӧ���������ɫӫ�⣬E12d�����ṹԭ���ɼ����Ա���;E16��E18d����ϸ����Ҫ�ֲ���CA����㼰��״�ؿ�����;P1��P3d������;P5��P14d����;P21d��3m������������ϸ��(ͼ2)��
����2.3�羵�۲���
����(1)ϸ����ֳ(ͼ3A��B):E12d��E16d�����ṹ�м�������ֳϸ����P1d��P7d�Կɼ�������ֳϸ��������Ϊ��Ⱦɫ�ʴֿ�״������ɢ�ң���Ĥ���壬�˷������;(2)ϸ������(ͼ3C��D):E16d��P1d��P7d���ɼ�����ϸ������P1d�Ӷ࣬���ֺ�Ⱦɫ��Ũ������ϸ����������E12dδ��������ϸ����
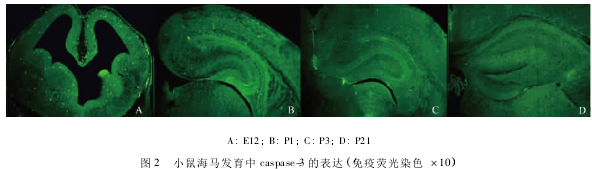
����2.4DCX��Caspase-3����ӡ�����������
����DCX��caspase-3����ӡ������������ͼ4�����ױ���������:(1)DCX���ױ�����:E18d��P3d������(P��0.01);P3��P5d�����Բ���(P��0.05);P7d��2m����(P��0.01);2��3m�����Բ���(P��d0.05)(ͼ5)��(2)Caspase-3���ױ�����:E18d��P3d������(P��0.01);P5��P21d����(P��0.01);P21d�����ȶ�(P��0.05)(ͼ5)��


����3������
��������������������������ͨ��ϸ����ֳ�������Э�����õ��ڵġ�DCX���о������ĺ��ʱ������ݱ�������ֳ��ǰ��ϸ����������ǿɷ�ӳ����ˮƽ�ߵ͡�Caspase-3��ϸ����������Ҫִ���ߣ�Ҳ���ж�ϸ��������������־(biochemicalmarker)����ϸ����ֳ���������̬ѧ�ϱ���ɿ��ķ����ǵ�������������
������ʵ���C57/BL6С����DCX�����黯Ⱦɫ�����ʾ:E12��E18d���ɼ���������ϸ������Walker�ȶ�С����̥���������о��������;P1��P14d����ϸ������������ų�һ���ܴ���P1d�����P14d��ʧ����ϯ�����ȶԴ�����о�������ƣ��Ʋ����Ϊ������Ԫ����ڣ����������״����Ǩ������;����ϸ���ֲ�:P1��P7d��Ҫλ�ڳ�״��(DG)������Ͷ��β㣬P14d��Ҫλ�ڿ�������1/2��P21d��Ҫλ���ǿ���ϸ���㣬P28d��3m�ǿ���ϸ��������ϸ�����٣����ֽ��������Muddanna�ȶԴ�����о�������ơ���ʵ��DCX����ӡ�����������ʾ:��̥��������2w��DCX���ױ���������������仯���ԣ��������ڽϸ�ˮƽ��2m�������ȶ�����Muddanna�ȶԴ�����о�������ƣ���ϱ�ʵ��羵�۲�����E12d��P1d��P7d���ɼ���ֳϸ�����Ʋ���̥������������Ϊ�����ṹϸ����ֳ�ֻ�Ǩ�Ƽ���������߷塣
�����ں����ṹ�������������ϻ�������ʼ��ͬʱ������ϸ����ֳ�������ͨ�����������̵�Э�����ò��ܱ�֤���������ķ������������ϻ����̡�Caspase-3��ϸ����������ø������Ӧ�ıؾ�֮·��Ҳ�ǵ����Ĺؼ�ø��ִ���ߡ�LIU�ȶԴ������о������ʾ:Casepase-3��P2d������ǿ���Ժ����ͣ�P21d�����ȶ��Ľϵ�ˮƽ��
������ʵ���C57/BL6С����Caspase-3����ӫ�⼰����ӡ�������������:E12d��P3dcaspase-3���Ա��������࣬P3��P21d�������٣�P21d�������ȶ�;��ϱ�ʵ��羵�۲����ɼ���P1����ϸ��������࣬�Ʋ�С�����ṹϸ�������߷���P3d���ҡ�
����������������ʵ��Ӧ��������֯��ѧȾɫ������ӫ������羵������C57/BL6С������������������ϸ����ֳ������ı仯���ɽ�����ϵͳ�۲��о��������ʾ:��̥������������ΪС����ϸ����ֳ�ͷֻ�Ǩ�Ƹ߷壬�����߷�λ��P1��3d;ͬʱ��������ӡ�������Է����е�ϸ����ֳ����������˶Ա��Զ�����⣬������֣�С�������������У���E18d��P5dDCX����ȱ�ֵ����caspase-3����ȱ�ֵ������p7��p3m�������������ǰ�ߣ��������������ʽ���������ͼ��أ�˵������������ֳϸ���������Զ��ڵ���ϸ�������Ժ����������������ʽ��������ࡣ�о����Ϊ����̽�ֺ����������������Լ������������Լ����Ļ����о��ṩ�˻����Ķ�����̬ѧ���ݡ�





