植物病理学论文第八篇:植物的免疫系统与诱导抗性研究
摘要:植物在生长过程中具有自己特殊的免疫体系来抗御病原生物的侵染,而且这种防御功能还可以通过调控来得以强化.植物免疫系统是植物防御生物及非生物伤害的生物化学屏障,是植物健康维护的重要基础.本文系统介绍了植物免疫及免疫调控的概念,分析了植物免疫及免疫调控的机制,提出了植物医学中免疫调控的相关策略,展望了免疫调控技术在植物医学中的发展趋势与方向,对丰富植物医学理论和实践具有重要意义和价值.
关键词:植物健康; 植物免疫; 信号途径; 免疫调控;
Immunoregulation——A New Concept of Phytomedicine
DING Wei LIU Ying
School of Plant Protection,Southwest University
Abstract:Plants have evolved their own innate immune system to resist the infection of pathogens,which is different from the immune system of animals.Moreover,this defense function can be reinforced through regulation.The plant immune system is a biochemical barrier against biotic and abiotic infections,which is an important basis for maintaining plant health.Focusing on the concepts of plant immunity and immunoregulation,this paper analyzes the mechanism of immune regulation,proposes relevant strategies for controlling the immune system of plants,and points out the development trend and direction of immunoregulation in phytomedicine.This should be of significance and value for enriching the theory and practice of phytomedicine.
1 植物免疫与免疫调控的概念
植物免疫及其机制一直是人们关注的重要学术领域.植物的生长发育以及自身结构的特殊性决定了植物免疫系统和动物免疫系统有明显的差异,人们对植物免疫和植物免疫诱抗剂乃至植物疫苗的概念一直存在着模糊的认识,这直接影响到了植物保护的实践活动.19世纪以来,研究者们逐渐意识到植物病害是由微生物引起的,但有的植物不易感病,且具有抗病性;1955年,Flor基于亚麻锈病的发生与植物抗病基因缺失的相关性,提出了“基因对基因”假说[1],明确了寄主植物基因不亲和识别病原生物是产生抗病性的重要机理.随后,众多与抗病相关的基因在植物中被发掘,说明植物对病害的抵抗力是受植物基因控制的,而且可以诱导表达,由此,植物免疫的概念逐渐产生.植物免疫(plant immunization)即由植物自身决定的其对病害的抗御能力,也称为植物的抗病性.这种抗病性是可以遗传的,可以在一定时期内自然表达,也可以在一定物质的诱导下表达.植物免疫常常是系统的,在对某种病原因子产生抗性后,还会表现出对其他因子的抗性和适应性.
随后,越来越多的研究结果表明,植物除了具有抵御外源物质侵袭的物理屏障(如角质层等)外,其内在还有一套复杂的“免疫系统”来识别和抑制病原物的侵染或发展.Jones等[2]于2006年总结了植物与病原物之间互作的两层防御系统,包括由病原菌相关分子模式触发的植物基础抗性PTI(PAMP-triggered immunity),以及病原效应子激发的植物免疫ETI(effector-triggered immunity).这些研究结果逐步完善了人们对植物天然免疫系统的认识.
当植物局部组织受到病原体或有益菌感染或定殖时,会在局部组织中引发植物自身的抗性,而且这种抗性可进行远源传递,即在远离感染或定殖部位的组织中也会引发诱导抗性,使植物获得增强的系统防御能力,这种诱导的系统抗性被称为系统获得性抗性(Systemic acquired resistance,SAR)或诱导系统抗性(induced systemic resistance,ISR)[3,4].此外,植食性昆虫和特定化学物质也会诱发抗性.植物诱导抗性为保护植物免受病原物危害提供了巨大的调控潜力,植物免疫诱抗剂也应运而生.
植物医学的免疫调控(immunoregulation),是指采用外源诱导的方法,调控植物自身内在的免疫系统,提高植物抵抗病原菌侵染的能力,达到维护植物健康的目的.这包括利用物理或者化学因子(免疫诱导子)诱导一些基因的表达,从而获得后天的诱导性抗性;也可以利用病原物的弱毒株系(免疫诱导菌)在植物活体内产生抑制强致病株系的侵染与发病等.在植物医学这一领域中,为了维护植物健康,免疫调控是最重要、最基础的保护途径之一.免疫调控,既可以通过对植物内在的天然免疫系统进行调控,让植物提高抵抗外源病原物的防御能力,也可以对植物生存的土壤环境进行调控,让土壤形成第二个强有力的免疫系统,共同维护植物的健康[5].
植物的免疫调控通常具有广谱性和系统性,一种免疫诱导物质作用于植物后,植物不仅会表现出对一些病原菌有抵抗力,通常还可能会对一些不良的环境条件产生抵抗力,这是植物诱抗剂有别于疫苗的重要特征.鉴于植物防御系统的复杂性,目前还几乎没有针对某一种病原菌的特异性疫苗,因此,将植物的免疫诱抗剂称为植物疫苗是不准确的.
2 植物的免疫系统
植物在生长过程中面临诸多威胁与挑战,例如,致病细菌可通过气孔、水孔或伤口进入植物后在植物细胞间隙(质外体)中增殖引起植物发病;真菌可以直接进入植物表皮细胞,或扩展菌丝在植物细胞之上、之间,甚至直达细胞;线虫和蚜虫以口针直接刺入植物细胞中进行取食.而在长期的协同进化过程中,植物也形成了自身的“防卫系统”来对抗环境中的各种不利因子.植物抵抗外界致病因子的系统可分3个屏障,分别为物理屏障、化学屏障和生物屏障[6].植物免疫实际上是植物化学屏障的重要组成部分.
在长期的进化过程中,植物形成了复杂的基因表达调控网络来保护自己,调节生长发育和对逆境的适应.在应对外界生物与非生物胁迫过程中,植物进化出了一系列复杂的免疫系统.如前所述,植物具有两种策略来探测和抵抗病原体,分别为病原物相关分子模式触发的免疫反应PTI和效应子触发的免疫反应ETI[2].
2.1 植物对于病原物的感应
植物对病原物的感应可分为两类.第一种感应是在寄主细胞的外部由病原相关分子模式(microbial-or pathogen-associated molecular patterns,MAMPs or PAMPs)的保守微生物激发子触发的,它能够被模式识别受体(pattern recognition receptors,PRRs)蛋白识别.病原相关分子模式是这个类别病原菌典型的基本组件,例如细菌鞭毛蛋白或真菌甲壳素等.植物对入侵病原体释放的内生分子也有反应,例如病原菌细胞壁或被称作危险相关分子模式(DAMPs)的角质层碎片.模式识别受体(PRRs)的刺激会引发病原物相关分子模式触发的免疫反应(PTI).
第二种感应则是涉及到胞内受体对病原体毒性分子(效应子,effectors)的识别,这种识别会引起效应子触发的免疫反应,该模式会引起植物和病原体之间的协同进化.与PAMPs完全不同,效应子是可变的且是非必要的,这种识别主要依靠多态的NB-LRR蛋白质产物,他们由大多数R基因编码,以特征核苷酸结合位点(nucleotide binding,NB)和富含亮氨酸重复序列(leucine rich repeat,LRR)结构域来命名,而效应子就是被NB-LRR蛋白识别,并激活相似的防卫反应(ETI).NB-LRR调节的抗病性能有效对抗只能生活在活体寄主组织(专性活体寄生)中的病原菌或半活体寄生病原菌,但不抵抗在定殖过程中杀死寄主组织的病原菌(死体寄生).
这两种感应是植物与病原体协同进化的结果.当病原菌突破寄主植物的第一道防线时,植物会通过感应病原体的MAMPs或PAMPs来触发自身的PTI反应,以此来抵抗病原物的侵染(图1所示第1阶段);但这种抵抗的程度并不是很高,仅能抵抗一部分病原物,很多病原物能克服植物的这一抗性,它们的策略就是分泌各种毒性效应子,从而干扰植物的基础抗性PTI,导致植物感病(如图1所示第2阶段),该过程被称为效应子激发的感病性(effector-triggered susceptibility,ETS).同时,植物也进化出了相应的应对机制,即产生专一性识别效应子的抗性基因(R基因)进行识别,从而触发强烈的ETI反应(如图1所示第3阶段,其中“Avr-R”即病原物的效应子被植物的NB-LRR蛋白所识别的过程).而在进化过程中,有的病原物会“丢弃”能被识别的效应子,并产生新的效应子(如图1所示第4阶段,病原物产生新的效应子,再次抑制植物的ETI),这些效应子可以帮助病原物抑制植物的ETI免疫,从而使植物感病,同时,有的植物也进化出了能识别这些新的效应子的R蛋白,从而再次获得对病原物的抗性.这就是植物免疫系统有名的“Z”字形模型(图1).
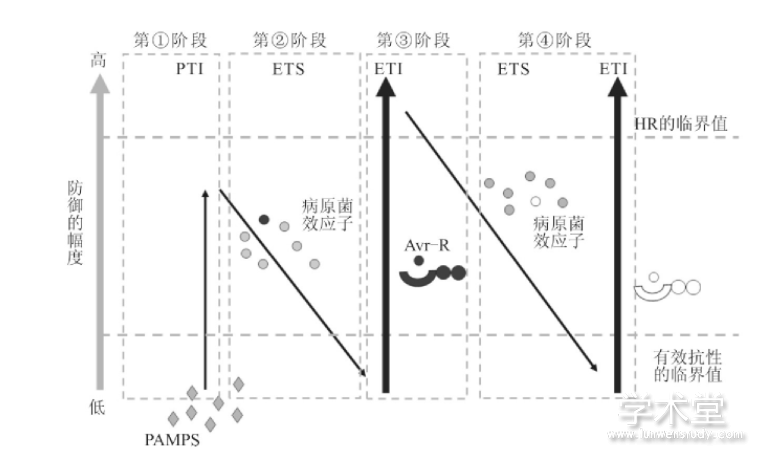
图1 植物免疫系统“Z”字形模型[2]
2.2 植物的过敏性坏死反应
植物与病原菌互作时,会存在两种情况,即亲和互作(compatible interaction)和不亲和互作(incompatible interaction).当病原菌侵染植物后,发生不亲和互作时,通常会引起侵染点周围局部组织或细胞的程序性死亡,这种现象叫做过敏性反应(hypersensitive response,HR)[7].
植物的过敏反应所涉及的程序性细胞死亡,不同于发育过程中的程序性细胞死亡,它与局部和全身防御反应的诱导具有关联性.过敏性细胞死亡通常通过病原体无毒基因产物与植物抗性基因产物之间的直接或间接相互作用来控制,并且它可以是多种信号传导途径的结果.离子流和活性氧物质的产生通常先于细胞死亡,但后者直接参与并随植物-病原体组合而变化[8].蛋白质合成,完整的肌动蛋白细胞骨架和水杨酸似乎也是细胞死亡诱导所必需的.细胞学研究表明,除了细胞内容物的实际模式和顺序在植物-寄生虫系统中不同外,还可能存在半胱氨酸蛋白酶的普遍参与,HR中的细胞死亡似乎更像是对植物其他部分的信号,而不是作为直接防御机制.
HR反应是植物的免疫反应之一,能产生局部坏死从而阻止病原物在体内的生长、繁殖和扩展.HR发生的早期,植物细胞会发生电解质流失,磷脂代谢加快.无论是非寄主植物HR还是品种特异性HR,最初阶段都有大量活性氧(ROS)产生,它们参与细胞膜脂质过氧化、细胞壁木质化和蛋白质重聚,直接杀伤病原菌.除此之外,活性氧还参与信号传递,介入或启动植物的HR反应,与过敏反应相关的活性氧的生成可能有多个系统.
2.3 植物的信号通路在防卫反应中的作用
在植物抗病反应过程中,抗病信号必须由内源信号分子从受侵染部位传导至整株植物,引起相应的系统性抗性,因而内源信号分子在植物抗病信号传导途径中起重要作用.目前已经知道,水杨酸(salicylic acid,SA)、茉莉酸(jasmonic acid,JA)和乙烯(ethylene,ET)这3种激素在调节植物抵抗各种病原物、昆虫以及非生物胁迫如受伤等的防御反应中起到重要作用[9].此外,脱落酸(abscisic acid,ABA)、赤霉酸(gibberellic acid,GA)、活性氧(ROS)也是重要的信号分子[10,11,12].
2.3.1 水杨酸信号途径
SA是植物产生系统获得性抗性的重要诱导因子,SA途径是极其重要的抗病信号传导途径[13].SA在植物抵抗活体寄生、半活体寄生病原物的防御反应以及SAR的建立中都扮演了重要的角色[14].SA是植物抗病信号转导和SAR形成过程中必不可少的信号分子[15],并且外源应用会导致发病机制相关(pathogenesis related genes,PR)基因的感应并增强对较宽范围的病原体的抵抗力[9],在病原菌袭击的植物组织中SA水平会增加[16].
NPR1基因(Non-expressor of pathogenesis-related genes 1)是SA途径的关键基因,该基因过量表达,能够激活植物的多种抗病相关基因,提高抗病性[17].SA的积累使细胞内还原势增加,NPR1寡聚体中的半胱氨酸残基分子间二硫键被水解还原,形成NPR1单体,NPR1单体具完整的核定位序列,使其能够转移到细胞核内并与结合在PR-1基因启动子区的TGA类转录因子相互作用,调控PR基因的表达和SAR产生[18,19].
2.3.2 茉莉酸信号途径
作为植物激素和信号化合物,JA在植物生长发育以及抗胁迫过程中具有重要的调控作用[20].JA对植物抗病反应的调控主要包括3个方面:(1)启动植物抗病相关基因转录;(2)诱导植物生成生物碱和酚酸,直接对病原菌产生毒害作用;(3)诱导植物生理结构的变化.JA在茉莉酸羧基甲基转移酶的作用下可生成茉莉酸甲酯(MeJA),激活植物防御反应[21].F-box蛋白COI1(Coronatine insensitive1)是JA信号通路的关键调节因子.
防御反应中的JA响应的基因表达主要由转录因子JIN1/MYC2(Jasmonate insensitive 1/MYC2)介导[22].据报道,AP2/ERF(Apetala 2/Ethylene-responsive factor)家族的几个成员参与了JA调节的应激反应[23].ERF1、ERF2、ERF5和ERF6控制JA反应性标记基因PDF1.2(Plant defensin 1.2)的表达水平,并提供对坏死性病原体的抗性[11].
2.3.3 乙烯信号途径
ET是重要的植物激素之一,由甲硫氨酸衍生而来,具有挥发性.ET参与了植物的多个生理过程,它可被多种生物与非生物胁迫所诱导.
ERFs是应激反应中ET信号通路的主要下游调节因子[11].转录因子EIN3(Ethylene insensitive 3)被认为可诱导ERF1基因在ET反应中的表达并激活防御反应[24].ET信号的另一个正调节因子是EIN2.在没有ET的情况下,CTR1(Constitutive triple response)抑制了EIN2.一旦其受体ETR1(Ethylene response 1)感知到ET时,EIN2的抑制得到缓解,从而激活ET信号[25].ET可以拮抗或促进SA或JA途径,从而实现更有效的防御反应.
2.3.4 各信号途径相互串扰
植物抗性形成的各个信号途径并不会完全孤立的.例如,SA途径和JA/ET途径之间还存在着密切联系,两者相互抑制或增强.SA主要诱导酸性PR蛋白,JA/ET主要诱导碱性PR蛋白;在成熟的烟草叶片中,所有碱性PR蛋白都被SA抑制,而所有酸性PR蛋白都被JA抑制;这说明SA途径和JA途径之间存在拮抗作用.SA途径和JA/ET途径之间的对话,是通过信号转导网络中的“节点”基因实现的.NPR1基因是研究较深入的“节点”基因之一[19].
3 植物诱导抗性
3.1 系统获得性抗性和诱导系统抗性
局部感染组织中PTI和ETI的激活通常在远离感染部位的组织中引发诱导的抗性,并且涉及一种或多种长距离信号,并在仍未受损的植物部分中传播增强其防御能力,这种病原体诱导的系统抗性被称为系统获得性抗性[3].PTI和ETI被迅速激活并在局部采取行动以限制特定入侵者在感染部位的生长,而SAR虽然需要更多时间来产生,但其具有强大的防御能力,通常可有效对抗病原体,且具有广谱性.植物中SAR赋予的免疫“记忆”可持续数周至数月,甚至可能持续整个生长季节.
在植物全身组织中,SAR的一个重要特征在于水杨酸(SA)水平的增加,这是SAR的标志之一.SAR的另一个标志是PR(Pathogenesis-related)基因的协调激活,其中有几个基因编码PR蛋白抗菌活性[26].PR-1是最具特征的PR基因之一,并且在许多植物中被当作SAR的标志.
在激活SAR后,SA信号被氧化还原蛋白NPR1转导,作为大量PR基因的转录共激活因子起作用.在未刺激的细胞中,NPR1通过分子间二硫键作为寡聚体在细胞质中隔离.在SA积累时,细胞氧化还原状态的变化介导NPR1的单体化,使得NPR1易位到细胞核中.在细胞核中,NPR1与TGA转录因子相互作用,并与WRKY转录因子一起激活SA响应性PR基因.NPR1要发挥正常功能还需要蛋白质被蛋白酶分解.同时,NPR1及其旁系同源物NPR3和NPR4被鉴定为具有不同亲和力的SA结合受体,从而影响NPR1的稳定性.
除病原体感染外,有益微生物,如植物根际促生细菌(plant growth-promoting rhizobacteria,PGPR)和非致病的植物根际促生真菌(plant growth-promoting fungi,PGPF),对植物根系的定殖也已被证明能刺激植物免疫系统,导致表型相似的广谱抗病性,通常也被称为诱导系统抗性[4].研究表明,大部分根际促生细菌介导的诱导系统抗性与病原体诱导的系统获得性抗性受不同信号传导途径的调节.
此外,植食性昆虫和特定化学物质也会诱发植物产生抗性.经过几十年的研究,人们逐渐认识到不同形式的诱导抗性受到互联信号通路的复杂网络的调节,其中植物激素起着重要的调节作用[16].由病原体、有益微生物和昆虫触发的诱导抗性信号传导途径部分重叠并共享共同的信号传导组分.这为植物提供了巨大的调控潜力,可以快速适应其生物环境,并以有效的方式利用其有限的资源促进生长和生存.
3.2 免疫诱抗剂
植物免疫诱抗剂是一类能够诱导植物免疫系统从而使植物获得或提高抵抗病菌能力的药物或微生物制剂,一般可分为植物免疫诱导子和植物免疫诱导菌[27].植物免疫诱导剂从作用机理上有别于动物的疫苗,但从功能上讲可以看作是植物的疫苗,因为动物疫苗是特异性的,而植物免疫诱抗剂常常是广谱的.
3.2.1 植物免疫诱导子
植物免疫诱导子是能诱导寄主植物产生免疫抗性反应的活性分子的总称,包括蛋白、寡糖、生物代谢产物或有机活性小分子.目前市面上常用的活性物质包括水杨酸(SA)、2,6-二氯异烟酸(INA)、苯丙噻二唑(BTH)、茉莉酸(JA)及其甲酯(MeJA)、β-氨基丁酸(BABA)等.
此外,蛋白质类以及糖类免疫诱导剂也是重要的免疫诱抗剂,如寡链蛋白、氨基寡糖素等在提高植物抗性,绿色防控病害中起到重要作用.
蛋白质类免疫诱导剂是一类能激活植物抗病性增强的蛋白质药物,多是从微生物中分离纯化的.据报道,目前已从极细链格孢、稻瘟病菌、大丽轮枝菌、灰葡萄孢菌等病原真菌以及侧孢短芽胞杆菌、解淀粉芽胞杆菌等生防细菌中分离获得了多个植物免疫诱抗蛋白,为后续蛋白质生物农药的开发提供了重要基础支撑[27].
糖类免疫诱导剂以寡糖类物质研究较多,其中,登记的该类生物农药中壳寡糖占据重要地位.壳聚糖存在于某些病原菌的细胞壁上,在植物与病原菌互作过程中,其降解产生的糖类片段(壳寡糖)可调控植物免疫产生抗性[28].因其原料广泛、易于降解等多种特点,目前已有大量以壳聚糖及其衍生物为原料的产品出现.
3.2.2 植物免疫诱导菌
植物免疫诱导菌是能激发植物自身免疫从而使植物获得抗病性或抗逆性的一类微生物.因其数量庞大,这类微生物是具有重要潜力的资源,不仅可以开发生防药剂,还可以用于探索植物免疫诱导剂.植物免疫诱抗菌在植物防御的生物屏障中发挥了重要作用.木霉菌是目前研究较多的植物免疫诱导菌之一,广泛存在于土壤中,是重要的生防微生物;随着研究的不断深入,目前已证实木霉菌在诱导植物免疫方面具有重要价值[29].
4 免疫调控技术在维护植物健康中的应用
免疫调控的核心是以物质或外源微生物来激活植物内在的抵抗机制,由外到内保障植物的健康.免疫调控技术被广泛应用在植物病毒病的防治上,同时对细菌性青枯病、番茄灰霉病等也有一定的辅助防治效果.
氨基寡糖素作为寡糖类诱导剂,对病毒病具有较好防效.防治烟草、辣椒、番茄病毒病的防效分别可达77.9%,77.0%和74.45%[30].除了具有防病作用外,该药还能提高作物产量,刺激作物的生长,是对经济作物无公害生产具有积极作用的生物农药.
寡糖·链蛋白是最近才登记应用的抗病毒蛋白质生物农药,可诱导植物产生系统抗性从而对病毒病具有较好的防效.田间应用表明,6%寡糖·链蛋白可湿性粉剂防治水稻条纹叶枯病、黄化曲叶病毒病、烟草花叶病毒病的防效分别为65%,68%和75%,具有良好的应用前景.
水杨酸及其类似物如BTH和INA也是具有广泛应用前景的诱导剂.BTH与水杨酸相比,其诱导效率更高,且在有效诱导浓度范围内对植株没有生理毒性或所产生的生理毒性难以从表象观察到.INA能诱导烟草、黄瓜以及拟南芥体内PR蛋白基因表达,激活β-1,3葡聚糖酶、6-磷酸葡糖苷脱氢酶、几丁质酶的活性.INA可处理植物整株、叶片或种子等,均能有效激活抗病防御反应,保护作物免受病毒、细菌和真菌等病原物的侵害.
虽然免疫诱抗技术已经得到了广泛应用,但由于受植物免疫程度、环境条件、药剂的特性等影响,免疫诱抗技术的效果还存在一定的争议,在实际应用中,还应注意以下几点:
(1)注重土壤免疫的调控,为植物的健康生长打下基础.坚持植物材料还田,增施有机肥,保障土壤微生物的活力;注意土壤的酸碱度变化,保持土壤正常的pH值;在作物种植过程中添加一些有益菌;施用土壤调理剂等[5].
(2)在作物生长的关键时期施用植物免疫诱抗剂.诱导植物产生自身的抗性有一定的过程,而且植物的诱导系统抗性具有持久性和广谱性,故应在早期施用诱抗剂,即在病原物初侵染之前施用诱导效果最好.一旦开始发病,诱抗的效果会大大降低.
(3)植物免疫诱抗剂的用量一定要适宜,使用要精准.一般免疫激发子在微量的情况下能更好地激活植物的抗性,过量易造成负效应,抑制植物的生长.
(4)选用具有诱抗效果的拮抗微生物或有益微生物效果更佳.施用有益微生物来形成生物屏障,是保护植物的重要一环[6].当施用的微生物同时能诱导植物产生抗性,更能有效地抵御生物因子和非生物因子对植物的伤害.
5 展望
免疫调控是一种绿色、安全的方法,该技术通过构建植物健康的生存土壤环境以及激发植物自身的免疫反应达到防控有害生物的目的.传统意义上的免疫调控仅仅局限于植物免疫诱抗剂,因植物诱导抗性是一种广谱性的防御反应,虽然能在一定程度上抵御病原物的侵染与为害,但在很多毁灭性病害如茄科植物青枯病、马铃薯晚疫病等面前仍然会束手无策.因此,在植物医学的未来研究中,免疫调控还有待新突破,如何使植物获得具有特异抵抗某种或某些病原物的能力是未来需要解决的重大科学问题.未来可关注以下3个方面:第一,发掘更多具有特异性识别病原物的R蛋白,植物的大多数R-基因蛋白是专门为识别病原菌效应子而进化的,针对不同的植物对病原物识别的差异,可探究更有效的R蛋白,为今后利用植物免疫系统防控病害提供新方法;第二,探索特异的代谢途径,为植物装配能分泌特定代谢产物来达到抑制特定病原物效果的相关代谢模块,以此来维护植物的健康生长;第三,研究植物根际和叶际微生物与植物抗性的关系,借助于代谢组学和合成生物学技术开发出更具有免疫调控价值的天然活性化合物.为了实现突破,应多学科交叉,注重新技术的运用.目前新兴的合成生物学具有巨大潜力,该学科的技术可允许我们直接探究物质的相关代谢通路与植物、微生物的互作,为免疫调控提供直接有效的支撑.
参考文献
[1]FLOR H H.Host-Parasite Interaction in Flax Rust-Its Genetics and Other Implications[J].Phytopathology,1955,45(12):680-685.
[2]JONES J D G,DANGL J L.The Plant Immune System[J].Nature,2006,444(7117):323-329.
[3]FU Z Q,DONG X N.Systemic Acquired Resistance:Turning Local Infection into Global Defense[J].Annual Review of Plant Biology,2013,64(1):839-863.
[4]PIETERSE C M J,ZAMIOUDIS C,BERENDSEN R L,et al.Induced Systemic Resistance by Beneficial Microbes[J].Annual Review of Phytopathology,2014,52(1):347-375.
[5]丁伟,李石力.植物医学的新概念---土壤免疫[J].植物医生,2019,32(2):1-7.
[6]丁伟,刘晓姣.植物医学的新概念---生物屏障[J].植物医生,2019,32(1):1-6.
[7]向婧姝.植物过敏性反应分子机制研究进展[J].江西农业学报,2018,30(12):41-46.
[8]HEATH M C.Hypersensitive Response-Related Death[J].Plant Molecular Biology,2000,44(3):321-334.
[9]BARI R,JONES J D G.Role of Plant Hormones in Plant Defence Responses[J].Plant Molecular Biology,2009,69(4):473-488.
[10]LAMB C,DIXON R A.The Oxidative Burst in Plant Disease Resistance[J].Annual Review of Plant Physiology and Plant Molecular Biology,1997,48(1):251-275.
[11]VERMA V,RAVINDRAN P,KUMAR P P.Plant Hormone-Mediated Regulation of Stress Responses[J].BMC Plant Biology,2016,16:86.
[12]WASZCZAK C,CARMODY M,KANGASJARVI J.Reactive Oxygen Species in Plant Signaling[J].Annual Review of Plant Biology,2018,69(1):209-236.
[13]AN C F,MOU Z L.Salicylic Acid and Its Function in Plant Immunity[J].Journal of Integrative Plant Biology,2011,53(6):412-428.
[14]DODDS P N,RATHJEN J P.Plant Immunity:Towards an Integrated View of Plant-Pathogen Interactions[J].Nature Reviews Genetics,2010,11(8):539-548.
[15]余朝阁,李天来,杜妍妍,等.植物诱导抗病信号传导途径[J].植物保护,2008,34(1):1-4.
[16]PIETERSE C M J,VANDER DOES D,ZAMIOUDIS C,et al.Hormonal Modulation of Plant Immunity[J].Annual Review of Cell and Developmental Biology,2012,28(1):489-521.
[17]张红志,蔡新忠.病程相关基因非表达子1(NPR1):植物抗病信号网络中的关键节点[J].生物工程学报,2005,21(4):511-515.
[18]SPOEL S H,KOORNNEEF A,CLAESSENS S M C,et al.NPR1 Modulates Cross-Talk between Salicylate-and Jasmonate-Dependent Defense Pathways through a Novel Function in the Cytosol[J].The Plant Cell,2003,15(3):760-770.
[19]VLOT A C,DEMPSEY D A,KLESSIG D F.Salicylic Acid,a Multifaceted Hormone to Combat Disease[J].Annual Review of Phytopathology,2009,47(1):177-206.
[20]WASTERNACK C,HAUSE B.Jasmonates:Biosynthesis,Perception,Signal Transduction and Action in Plant Stress Response,Growth and Development.an Update to the 2007Review in Annals of Botany[J].Annals of Botany,2013,111(6):1021-1058.
[21]SEO H S,SONG J T,CHEONG J J,et al.Jasmonic Acid Carboxyl Methyltransferase:A Key Enzyme for JasmonateRegulated Plant Responses[J].Proceedings of the National Academy of Sciences,2001,98(8):4788-4793.
[22]EULGEM T,SOMSSICH I E.Networks of WRKY Transcription Factors in Defense Signaling[J].Current Opinion in Plant Biology,2007,10(4):366-371.
[23]MIZOI J,SHINOZAKI K,YAMAGUCHI-SHINOZAKI K.AP2/ERF Family Transcription Factors in Plant Abiotic Stress Responses[J].Biochimica Et Biophysica Acta(BBA)-Gene Regulatory Mechanisms,2012,1819(2):86-96.
[24]SOLANO R,STEPANOVA A,CHAO Q,et al.Nuclear Events in Ethylene Signaling:ATranscriptional Cascade Mediated by ETHYLENE-INSENSITIVE3and ETHYLENE-RESPONSE-FACTOR1[J].Genes&Development,1998,12(23):3703-3714.
[25]JU C,YOON G M,SHEMANSKY J M,et al.CTR1Phosphorylates the Central Regulator EIN2to Control Ethylene Hormone Signaling from the ER Membrane to the Nucleus in Arabidopsis[J].Proceedings of the National Academy of Sciences,2012,109(47):19486-19491.
[26]VAN LOON L C,REP M,PIETERSE C M J.Significance of Inducible Defense-Related Proteins in Infected Plants[J].Annual Review of Phytopathology,2006,44(1):135-162.
[27]邱德文.我国植物免疫诱导技术的研究现状与趋势分析[J].植物保护,2016,42(5):10-14.
[28]王文霞,尹恒,赵小明,等.糖链植物疫苗研究新进展[J].中国植保导刊,2014,34(9):17-22.
[29]陈捷.木霉菌诱导植物抗病性研究新进展[J].中国生物防治学报,2015,31(5):733-741.
[30]赵小明,杜昱光,白雪芳.氨基寡糖素诱导作物抗病毒病药效试验[J].中国农学通报,2004,20(4):245-247.





