
����ϸ�����Ӽ�������㷺����������ϸ������ϵͳ,��� �о����� ϸ �� �� ��CCL-2��CX3CL-1��CCL-1��ϸ�����ӿ���ͨ�������С����ϸ������ʹ��������������,��ʹ�ı����Ŵ�ѧ�о������롣�����Ŵ�ѧ��ָ��DNA����û�иı������»����������Ŵ��ĸı�,�����鵰����������������Ҫ�ı����Ŵ����η�ʽ�������о������鵰��3������9λ(H3histone subunitat lysine residue 9,H3K9)�� �� �� �� �� ʹ �� �ء�
����CCR-8[chemokine(C-C motif)receptor 8]��Ϊϸ������CCL-1������������,�书������ʹ���ڹ�ϵ���о���δ���������ʱ��о�����С���п�ʹģ�͵Ļ�����,�۲켹��CCR-8�����Ŵ����ؼ����ʹ��������Ӱ�졣
���������뷽��
����
�����Լ��������߷���(����:11062431),�������(Sigma��˾,����),������(Sigma��˾,����),��H3K9(Sigma��˾,����)С��һ������CCR-8С��һ��(Sigma��˾,����)�ͼ�������ø�������С�����(SC-2034,Santa��˾,����),Von Frey��ά˿��Model 400��ʹ�̼���(IITC��˾,����)��
����ʵ�鶯����༶��������������С��64ֻ,����18~20g,��ʵ�鶯�������ṩ������С��ʵ��ǰ����1��,12h/12h��ҹ����,�¶�(23±1)��,���ɽ�ʳ��ˮ��ʵ�鶯��ʹ������ʵ�鶯����������ع涨��
����ʵ�����64ֻС���������Ϊ:������(C��)���п�ʹ��(INC��)���п�ʹ+�鵰��ȥ������ø���Ƽ���(INC+SAHA��)���п�ʹ+�鵰������ת��ø���Ƽ���(INC+ACA��),����ÿ��10ֻ������ʹ��Ϊѧ���,ÿ������6ֻ�������4���⼹��H3K9��CCR-8���ױ��
����С���п�ʹģ���Ʊ�С���߷�������������,��������Һ�צ�Ų�,���������ܵķ����Ʊ��п�ʹģ��:ʹ��������Ƭ�����������ֺ���������п�5mm�������п�,�п����Ƥ����,���ۿ�����������Ⲣ������Է���,��������ֹ���ŵ��������пڴ�ɴ��ѹ��ֹѪ,ʹ��6-0����˿������ʽ���,������ע3��λ��ù��Ԥ����Ⱦ��
����INC+SAHA���INC+ACA��С��ֱ�����ǰ1d����ǰ2h����������1~4dÿ��������Ϊѧ���Ժ�ǻ�ֱ�ע��50mg/kg��SAHA��5mg/kg��ACA,����ҩ�������ҩʱ��ο�����Ԥʵ������
����С����ʹ��Ϊѧ��¼��ǰ1d(T0)��2h(T1)������1d(T2)��2d(T3)��3d(T4)��4d(T5)��5d(T6)��6d(T7)��7d(T8)��14d(T9)С���е������ֵ(mechani-cal withdrawal threshold,MWT)�� �� �� �� DZ �� ��(paw withdrawal latency,PWL)��MWT���:���л�������������˿������,С�������л�������Ԥ��Ӧ30min,�Բ�ͬ���ȵ�Von Frey��ë�̼��������,������������д�0.2g��ʼ,����ë����������Ϊ��ȫ������,�����̼�2s,����5��,ÿ�����ټ��15s����3�β�̧��,����һ����������ë;��3��̧��,�ص�һ����������ë,ֱ��ÿ5�β�������3��̧�ȡ��ܹ�����3/5��̧�ȵ����VonFrey��ë�Ŀ�����ΪMWTֵ��
����PWL���:���л�����������3mm��IJ�������,С�������л�������Ԥ��Ӧ30min,���ȷ���̼�������С�����,���俪ʼ��С�����̧�Ȼر���ֹ,�ȴ̼�ǿ��������ʵ�������ά��һ��,�Զ��ж�ʱ��Ϊ25s,�����ⶨ5��,ÿ�μ��5����,ȡ��3��ƽ��ֵ��ΪPWLֵ��
����Western blot���С���� H3K9��CCR-8���ױ�����ͱ�����(30mg/kg)�������¶�ͷ����С��,���ϲ���ȡL4~L6������������,�ο����� �� ��Western blot�� �� �� �� С �� �� ��H3K9��CCR-8���ױ���(H3K9��CCR-8һ��Ũ��Ϊ1��500)������������Photoshop�������лҶȷ���,Ŀ�ĵ��������Ҷ�ֵ����Ӧβ-actin�Ҷ�ֵ֮�ȷ�ӦĿ�ĵ��ױ��
����ͳ�Ʒ�������SPSS 16.0ͳ��ѧ���������������Ծ���±����(��x±s)��ʾ����ʹ��Ϊѧָ�����˫�����ظ������������,��������Ƚϲ���Bonferroni t����,Western blot����ʹ�õ����ط������,��������Ƚϲ���q���顣
�������
����
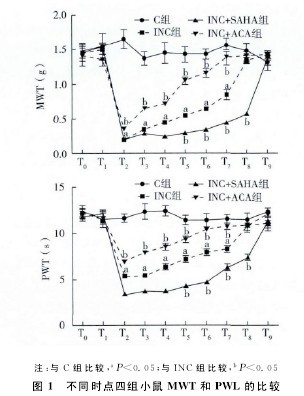
������T0ʱ�Ƚ�,T1~T9ʱC��MWT��PWL�Ƚϲ�����ͳ��ѧ���塣��C��Ƚ�,T2~T7ʱINC��MWT�� �� �� �͡�PWL�� �� �� �� (P <0.05)����ICN��Ƚ�,T5~T8ʱINC+SAHA��MWT���Խ��͡�PWL��������(P<0.05),T2~T7ʱINC+ACA��MWT�������ߡ�PWL�����ӳ�(P<0.05)(ͼ1)����ͼ1��
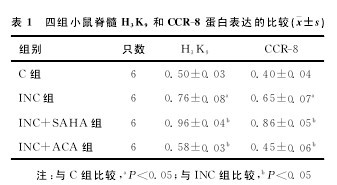
������C��Ƚ�,INC��H3K9��CCR-8���ױ�����������(P<0.05)����ICN��Ƚ�,INC+SA-HA��H3K9��CCR-8���ױ����������� (P<0.05);INC+ACA��H3K9��CCR-8���ױ������Լ���(P<0.05)(��1)������1��
��������
����������֯���˻���֢�������˺��Դ̼�������ǿ�ķ�Ӧ��������ʹ������,����Ӱ�컼�ߵ�����ָ�,�����Ŀǰ�Բ�ʮ��������о�����,С����ϸ����ʹ����������������Ҫ����,��ͨ���������ӡ�����ϸ�����ӡ���֢���ʵȼ�����������Ԫ��������,����ʹ����������,�������������ڸù��̵����� �� �� �� �� �� �� �ӡ�
����CCR-8�� Ϊ �� �� �� ��CCL-1���������������ʹ�������ķ�չ,�书�ܵ��ں�ʹ��������ϵ���о��в������
���������Ŵ�ѧ��ָDNA����ΧȾɫ������,�ڲ��ı�DNA���е������Ӱ�����ı���鵰��β�������Ṳ�����κ�DNA��������֪�����ֱ����Ŵ�������ʽ,���ֱ仯���Ƴ�����������ϸ������֯������Ӧ�Ըı��йء�Ŀǰ,�鵰�����������о�����鵰��������ʽ,ͨ���鵰������ת��ø(Histone acetyltransferases,HATs)���鵰��ȥ������ø(Histone deacetylases,HDACs)���ڻ���ƽ��:�鵰�����������Ⱦɫ���ɳ�,ת¼������ǿ;�鵰��ȥ���������DNA��������,����ת¼��Ĭ���鵰����������ȥ����������Ϊ�������Ͳ�����������������Ҫ������,���а���ʹ���������̡�ҩ��ѧ�о���ʾ,ʹ��HAT��HDAC���Ƽ���ϻ�̼��鵰����������Ӱ�������ʹģ�͵���ʹ��Ϊ�����о�Ҳ����ʹ��HDAC���Ƽ�SAHAʹС���п�ʹģ�������4~7��ʹ��������ǿ,HAT���Ƽ�ACA��ɻ���С���п�ʹģ�������2~6��ʹ����������һ���о����ָ�ǻ����ע��HDAC���Ƽ�SAHA�����鵰��H3K9��CCR-8������������,��HAT���Ƽ�ACA�����Լ��ټ����鵰��H3K9��CCR-8,��һ���Ҳ����ʹ��Ϊѧ�ı�һ�¡�
������������,����ˮƽ������������ı����Ŵ�ѧ���ػ������п�ʹʹ�����������з�����Ҫ����,�����ҩ���Ԥ��Ϊ�ٴ��п�ʹ�����ṩ�µ�˼·��





