
毛花猕猴桃(Actinidia eriantha Benth.)果肉翠绿色,风味较浓,具有较强的抗病性。目前商业栽培的猕猴桃种类主要是中华猕猴桃、美味猕猴桃(牛歆雨 等,2007),生产中应用的毛花猕猴桃品种极少。毛花猕猴桃绝大部分处于野生状态,主要分布于长江以南的广大丘陵地区,其中以江西、湖南、福建、广西等地的野生种质资源最为丰富。毛花猕猴桃是猕猴桃属植物中抗坏血酸(ascorbicacid,AsA)含量较高的种类之一,其果实中的 AsA 含量高达 5.00 ~ 13.79 mg · g-1FW,是中华猕猴桃的 3 ~ 4 倍(钟彩虹 等,2011)。由于具有这些特性,毛花猕猴桃具有广阔的开发利用前景。
近年来,国内外有关植物体内 AsA 的研究主要集中在抗氧化作用、光合保护、增强酶活性、促进细胞分裂、植物信号传导等方面的功能分析、AsA 合成途径与降解、转运与运输等生理代谢过程等方面。同时,在对植物抗坏血酸相关酶基因的研究上已在刺梨(安华明 等,2005)、柑橘(杨晓燕,2011)和猕猴桃(Laing et al.,2007;Bulley et al.,2009)等果树中广泛开展。而对于毛花猕猴桃的研究只侧重于 AsA 合成途径及相关基因的克隆等相关研究(Crowhurst et al.,2008;侯长明 等,2009)。有关野生毛花猕猴桃叶片和果实 AsA 含量与 SSR 标记的关联分析尚未见报道。
关联分析(association analysis,AA)作为在植物数量性状研究和植物育种中的最新开始应用的一种分析方法,它通过鉴定种质群体内性状与分子遗传标记或候选基因的关系,可以使 QTL 分析得到补充和提高,使目标性状精确定位并可广泛应用而不受杂交群体的限制(杨小红 等,2007)。根据扫描范围,关联分析可分为全基因组和候选基因两种途径(Flint et al.,2003)。全基因组关联分析基于遗传标记,可以对目标性状进行染色体定位;而候选基因关联分析则是对候选目的基因序列在种质间的差异进行研究,可以更加精确地鉴定优异等位基因。Hansen 等(2001)最早通过全基因组关联分析寻找出与野生甜菜抽薹基因紧密连锁的标记,加快了候选基因的定位进程。曹珂等(2012)在对桃的单果质量与6个物候期性状的遗传进行关联分析研究时得到了27个与桃单果质量及6个物候期性状关联的数量性状位点。文自翔等(2008)在对中国栽培和野生大豆农艺品质性状与 SSR 标记的关联分析中检测出栽培群体的大豆和野生大豆种质中分别有 27 个和 34 个位点与农艺形状性状相关。Breseghello 和 Sorrells(2006)利用 18 个 SSR 标记对 95 份小麦品种的籽粒性状进行了关联分析,结果发现 Xwmc111、Xgwm30 和 Xgwm261 与籽粒宽度存在显著相关。此外,全基因组关联分析还在其他一些作物上得到了广泛的应用,但在猕猴桃中尚未见报道。随着关联分析在分子遗传育种和基因组学研究中的不断深入,通过分子标记的开发、寻找候选基因及功能基因验证的研究,为进一步揭示数量性状的遗传机制及作物的遗传改良提供了新的途径。
在对江西省武功山境内野生毛花猕猴桃资源调查收集的 70 份野生种质叶片和果实的 AsA 含量变异分析的基础上(汤佳乐 等,2013),利用覆盖全基因组的 SSR 引物分析其遗传多态性,旨在为野生毛花猕猴桃资源的合理开发与利用奠定基础。同时运用一般线性模型(general linear model,GLM)和混合线性模型(mixed linear model,MLM)进行野生毛花猕猴桃 AsA 含量的关联分析,寻找与 AsA 含量相关的标记位点,为揭示野生毛花猕猴桃 AsA 生成的分子特征提供理论依据,以促进有效改良猕猴桃的果实性状与种质创新。
1、 材料与方法
1.1 材料及其基因组 DNA 的提取
2012 年 7 月中旬在江西省武功山境内随机采集 70 份野生状态下毛花猕猴桃种质的健康无损伤的成熟叶片,用冰盒保存并运回实验室。用 CTAB 法提取野生毛花猕猴桃幼嫩叶片的基因组 DNA,用紫外分光光度计检测 DNA 质量和浓度,–20 ℃保存备用。
1.2 SSR 分析
参照 Testolin 等(2001)用 SSR 构建了猕猴桃遗传连锁图,选用广泛分布在猕猴桃基因组中 70对 SSR 引物(Palombi & Damiano,2002;Zhen et al.,2004;Korkovelos et al.,2008),首先对 8 个AsA 含量差异较大的野生毛花猕猴桃种质材料进行扩增,筛选出多态性较好的 21 对 SSR 引物,再对 70 份野生毛花猕猴桃种质资源进行扩增。PCR 扩增程序为 94 ℃预变性 5 min;94 ℃变性 45 s,50 ~ 60 ℃退火 45 s,72 ℃延伸 45 s,30 个循环;72 ℃延伸 10 min。扩增产物用 10%聚丙烯酰胺凝胶电泳检测,快速银染法染色。
1.3 数据统计分析
利用 Microsoft Office Excel 对野生毛花猕猴桃成叶和幼果的 AsA 含量进行统计,根据丙烯酰胺凝胶电泳和银染检测的结果,将每条 SSR 引物扩增的每一条带视为 1 个位点,统计位点总数和多态性位点数多态性信息含量(PIC)等参数值,相同迁移率的记为 1,无条带的记为 0,不具多态性的条带不予统计。应用 Structure 2.3.1 软件进行群体遗传结构分析,并计算材料相应的群体结构 Q 值以确定 70 份野生毛花猕猴桃种质群体的遗传结构。先假定各个位点均是独立的,并估算最佳群体数(K),K 的取值范围为 2 ~ 10,将 MCMC(markov chain monte carlo)开始时的不作数迭代(lengthof burn-in period)设为 10 000 次,再将不作数迭代后的 MCMC 设为 10 000 次,然后选取一个合适的亲缘关系 K 值。
分别应用 Tassel 2.1 软件中 GLM 和 MLM 两种程序进行野生毛花猕猴桃 AsA 含量与 SSR 标记位点的关联分析。GLM 分析中以 Q 值作为协变量进行回归分析;MLM 采用群体结构 Q 值和亲缘关系 K 值相结合的方法进行分析,选择计算每个标记的遗传力(calculate heritability for each marker)的计算方式,分析方法选择 EM。利用 AsA 含量表型变异数据对标记逐一进行回归分析和关联作图,并计算标记对表型变异的解释率。
2、 结果与分析
2.1 SSR 多态性分析
利用 70 条广泛分布于猕猴桃基因组的 SSR 引物对野生毛花猕猴桃材料进行扩增,52 条引物能够扩增出条带,且每条引物能够扩增出 2 ~ 12 条带。再对 52 条 SSR 引物进行筛选,得到稳定性好、扩增性强和多态性好的 SSR 引物 21 条,一共扩增出 127 个多态性等位位点。通过对野生毛花猕猴桃基因组 PCR 扩增,多数 SSR 引物能够在野生毛花猕猴桃材料中表现出多态性,不同的 SSR 引物的扩增结果在总等位位点数、多态性位点数、等位位点亮度等方面都表现出了差异。在 70 份毛花猕猴桃材料中,21 条 SSR 引物扩增得到的基因片度长度大概在 75 ~ 400 bp 的变化范围,等位位点数目在 2 ~ 12 之间,平均每条 SSR 引物可以检测到 6.04 个等位位点,其中引物 722 检测到了 5 个等位位点变异(图 1)。利用 SSR 标记对野生毛花猕猴桃扩增得到的图谱,将扩增得到的多态性位点进行赋值进行关联分析。
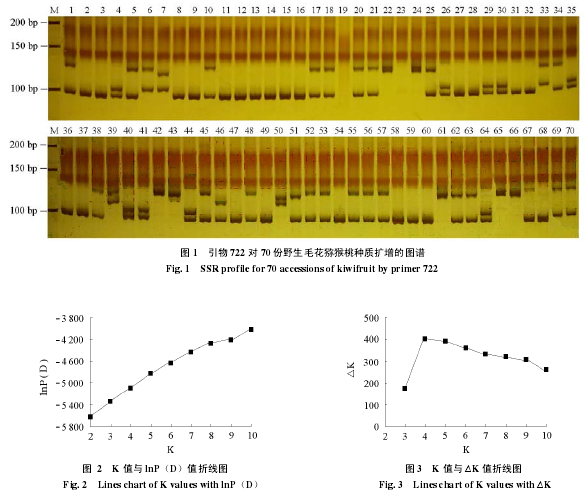
2.2 群体结构分析
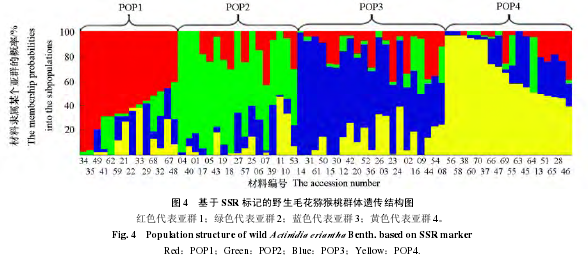
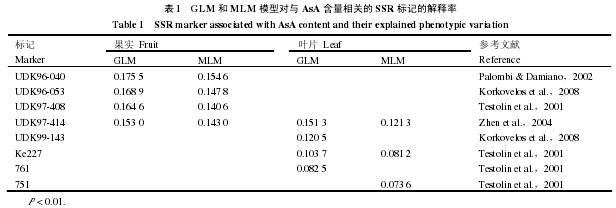
为了估测野生毛花猕猴桃种质群体的遗传结构,基于 SSR 标记的数据,利用 Structure 2.3.1 软件的等位基因发生频率相关模型进行群体结构分析。对 70 份野生毛花猕猴桃种质进行组群划分结果测试,重复 10 次,2 ~ 10 组群(K = 2,3,4,……,10),将测试过程中不同的等位变异频率特征类型数 K 给出的不同的 lnP(D)平均值(似然值),绘制成散点图(图 2)。基于数学模型的群体结构分析发现样本中的 lnP(D)随着 K 值得增加呈持续增大。因此,最佳的群体结构数必须通过 ΔK 来确定。由图 3 可以看出,ΔK 在 K = 4 出现拐点。根据 Evanno 等(2005)的描述,可以判断 70 份野生毛花猕猴桃群体可被分为 4 个亚群(图 4)。这 4 个亚群分别包含 14、17、21 和 18 份材料。
2.3 SSR 标记与 AsA 含量的关联分析
从前期研究结果(汤佳乐 等,2013)中发现,70 份野生状态下的毛花猕猴桃桃种质资源不同植株的成熟叶片和幼嫩果实 AsA 含量存在丰富的变异,变异系数分别为 41.30%和 21.88%,反映了野生毛花猕猴桃种质中存在较大的的遗传差异。本研究中则是基于对野生毛花猕猴桃准确测定成熟叶片和幼嫩果实的基础上,经过 SSR 多态性分析和群体遗传结构分析,将野生毛花猕猴桃群体分为4 个亚群,应用 Tassel 2.1 软件中的 GLM 模型和 MLM 模型两种程序进行野生毛花猕猴桃叶片和果实 AsA 含量与 SSR 标记的关联分析。
在 GLM 模型中,以 4 个亚群中的各个材料所对应的群体结构 Q 值作为协变量,将 SSR 标记与AsA 含量的变异进行回归分析,寻找与 AsA 含量相关联的标记并确定其解释率;在 MLM 模型中,采用群体结构 Q 值和亲缘关系 K 值相结合的方法进行回归分析,并确定其相关联标记的解释率。
GLM 模型分析结果(表 1)显示,在 P < 0.01 水平上,所检测的 21 对 SSR 引物的 127 个多态性标记位点中,有 7 个 SSR 标记位点与叶片和果实的 AsA 含量相关。有 4 个标记位点与叶片的 AsA含量相关,4 个与果实的 AsA 含量相关,其中标记位点 UDK97-414 同时与叶片的 AsA 含量、果实的 AsA 含量相关。各个 SSR 标记位点对叶片和果实 AsA 含量的变异的解释率在 0.082 5 ~ 0.175 5之间,解释率最大的标记位点是 UDK96-040,为 0.175 5,与果实的 AsA 含量相关,其次是标记位点 UDK96-053,解释率为 0.168 9;而解释率最小的标记位点是 761,解释率为 0.082 5,与叶片的AsA 含量相关。
MLM 模型分析的结果(表 1)显示,在 P < 0.01 的水平上,共有 6 个 SSR 标记位点分别与叶片和果实 AsA 含量相关,比 GLM 模型分析结果中的少 1 个,其中有 5 个 SSR 标记位点与 GLM 模型分析所得到的相同,标记位点 751 只有在 MLM 模型分析中检测到,与叶片的 AsA 含量相关的解释率为 0.073 6。标记位点 UDK97-414 同时与叶片和果实的 AsA 含量相关。MLM 模型中各个标记位点的解释率在 0.076 3 ~ 0.154 6 之间,低于 GLM 模型分析得到的解释率。
3、 讨论
3.1 野生毛花猕猴桃 SSR 多态性及群体结构分析
AsA 含量作为衡量果实品质中一个重要的指标,选育出高果实 AsA 含量的品种一直是育种工作者的目标之一。因此分析毛花猕猴桃 AsA 含量的多样性以及基于 SSR 分子标记的多态性分析,对于猕猴桃育种工作具有重要的指导意义。本研究中以 AsA 含量变异丰富的野生毛花猕猴桃种质为材料,在前期进行叶片和果实 AsA 含量变异分析的基础上(汤佳乐 等,2013),利用 SSR 分子标记进行多态性分析,21 条多态性高的 SSR 引物一共检测出 127 个等位变异位点,扩增得到的基因片度长度大概在 75 ~ 400 bp 的变化范围,变异范围在 2 ~ 12 之间,平均每条 SSR 引物可检测到 6.04个等位位点。可见,野生毛花猕猴桃不仅在叶片和果实 AsA 含量上具有丰富的多样性,而且基于SSR 分子标记在对野生毛花猕猴桃种质材料的识别和区分上也具有多态性高的特点。野生毛花猕猴桃叶片和果实 AsA 含量变异的多样性及基于 SSR 的多态性分析结果表明,本研究所选择的资源群体适合于开展野生毛花猕猴桃 AsA 含量的关联分析。通过对野生毛花猕猴桃种质材料的叶片和果实AsA 含量与 SSR 标记的关联分析,将有助于深入发掘并充分利用高 AsA 含量的基因资源,加速高AsA 含量基因的研究和育种改良。
对供试种质资源进行遗传多样性和群体遗传结构评价,是进行关联分析的前提和基础(Harris &Stokesbury,2010)。本研究中使用 STRUCTURE 软件对供试野生毛花猕猴桃种质进行的基于 SSR标记的群体结构分析,70 份供试种质可以分为 4 个亚群,分别包含 14、17、21 和 18 份材料。群体分析的结果分析是通过计算各个供试种质相应的 Q 值,进行基于数学模型的聚类划分,通过将野生毛花猕猴桃群体中各个种质的所得到相应的 Q 值作为协变量纳入回归分析中,校正了 70 份种质材料的群体结构,较大程度上减少群体结构对关联分析的影响,降低供试种质群体结构引起的伪关联的概率,进而有效提高了关联分析的准确性。
3.2 野生毛花猕猴桃 SSR 标记的关联分析
近年来,关联分析已成功应用于植物研究。顾竟等(2011)采用 GLM 一般线性模型,利用 59个 SSR 标记对豌豆的 19 个形态性状进行关联分析,结果显示 SSR 位点间有较高的多态性和一定程度的连锁不平衡,一共检测出 32 个 SSR 标记位点与 14 个表型性状相关联,其中有一些 SSR 标记与两个或多个形态性状相关联。Ehrenreich 等(2007)对来自中欧的 96 份拟南芥种质群体进行分析,并使用 MLM 混合线性模型的关联作图方法鉴定出 3 个与分枝变异显著关联的位点。Zhang 等(2010)和 Price 等(2010)认为在进行全基因组关联分析中 MLM 模型比 GLM 模型更适合。赖勇等(2013)对 113 份大麦亲本材料进行了 SSR 遗传多样性分析及群体遗传结构分析,结果发现使用 MLM 模型鉴定的显著关联位点要明显比 GLM 模型鉴定的少 3 个,两种模型分析中分别寻找到了 9 个与株高、穗长、芒长、穗粒数相关联,6 个与株高、芒长和小穗着生密度相关联的标记。
本研究中应用 Tassel 2.1 软件中的GLM 和MLM 两种模型分析方法进行野生毛花猕猴桃 AsA含量的关联分析,分别找到了 7 个和 6 个与果实、叶片相关的标记,其中 MLM 分析检测到的 5 个 SSR标记同样也在 GLM 分析中所检测到,标记 751 与叶片的 AsA 含量有关,只有在 MLM 模型分析中检测到;GLM 模型分析检测到与叶片的 AsA 含量有关的标记 761,却在 MLM 模型分析中未检测到。
MLM 模型分析结果比 GLM 模型分析结果要少 1 个,主要原因是 GLM 分析中只考虑群体结构 Q 值对关联分析的影响,而 MLM 分析中不仅考虑了 Q 值,还考虑到了亲缘关系 K 值对关联分析的影响,提高了关联分析的准确性,使结果更加准确可靠。
4、 结论
本研究中解析了野生毛花猕猴桃 AsA 含量关联位点内各等位变异的表型效应,在 AsA 含量的表型数据和分子标记检测得到的基因型数据之间用特定的等位变异建立起联系,发掘了与果实及叶片 AsA 含量关联的优异标记 SSR 位点 UDK96-040、UDK96-053、UDK97-408、UDK97-414 和 Ke227等。





