
植物竞争是群落中普遍存在的现象, 是影响森林生态系统的重要因素, 也是生态学研究植物生长、 种群动态和群落演替方向的核心问题之一。 竞争的结果不仅影响个体的生存、 生长和繁殖,而且影响种群空间分布、 动态和群落的物种多样性。 对个体竞争进行数量化研究, 能更准确地反映个体间的竞争状况, 进而解释和预测整个群落的动态变化过程。 近年来关于常绿阔叶林、 针叶林和针阔混交林等的优势种群的竞争已有不少报道, 而对落叶阔叶林中的优势种群竞争研究较少。 进一步研究落叶阔叶林中的优势种群竞争, 对揭示落叶阔叶林种群生态特性、 群落形成及其稳定性与演替规律等都具有重要意义。 龙王山地处中热带北缘—暖温带南缘的过渡性气候类型, 有着极其特殊的地理位置, 是中国沿海继泰山以外的另一个直接承受北方吹来的寒流的山峰, 分布在海拔约900~1 500 m 的落叶阔叶林是龙王山森林植被的主体和核心, 分布面积大, 林相保存完整, 种类组成复杂,优势树种多, 群系类型多具典型性和独特性。 因此, 笔者研究龙王山落叶阔叶林优势树种的种内种间竞争状况, 旨在为深入研究亚热带落叶阔叶林中的优势种群结构及演替进程, 以期为落叶阔叶林的保护及可持续利用提供科学依据。
1、 研究区自然概况
龙王山自然保护区位于浙江省安吉县, 其东南与浙江省临安市天目山区的西天目乡接壤, 西南与安徽宁国县相邻, 北连该县章村镇, 东北接该县报福镇。 区域地理位置为 30°22′30″~30°27′30″N, 119°22′30″~119°26′15″E, 主峰海拔为 1 587.4 m, 是浙北最高峰。 山体坡度较大, 岩石垂直节理发育, 多悬崖陡壁, 河谷深切, 峰谷交错, 地形极为复杂。 小气候类型复杂, 气候垂直变化明显。 山麓年均温 15.5℃, 无霜期 225 d, 年降水量约 1 640 mm, 集中在 6-7 月, 属于亚热带季风气候。 龙王山岩石以凝灰岩为主, 另有少量流纹岩。 主要的土壤类型有红壤、 山地黄壤、 山地黄棕壤、 山地草甸土, 以及高位沼泽。 土壤腐殖质层较厚, 肥力较高。 适宜的气候, 丰富的小气候类型, 复杂的地形和肥沃的土壤, 为植物生长提供了优越的自然条件。
试验地植物种类较丰富。 调查数据显示, 胸径 5 cm 以上的树种有 71 种, 主要有小叶白辛树Pterostyrax corymbosus, 四照花 Cornus kousa, 毛山荆子 Malus baccata, 茅栗 Castanea seguinii, 灯台树Cornus controversa, 玉铃花 Styrax obassia, 米心水青冈 Fagus engleriana, 黄山栎 Quercus stewardii, 青钱柳 Cyclocarya paliurus, 华山矾 Symplocos chinensis, 锥栗 Castanea henryi, 杜梨 Pyrus betulaefolia 等。 灌木层中主要有垂丝卫矛 Euonymus oxyphyllus, 合轴荚蒾 Viburnum sympodiale, 中国绣球 Hydrangea chi-nensis, 宜昌荚蒾 Viburnum erosum, 伞花石楠 Photinia subymbellata, 西南卫矛 Euonymus hamiltonianus,下江忍冬 Lonicera modesta, 云锦杜鹃 Rhododendron fortunei 等。 草本层常见植物有书带薹草 Carex roche-brunii, 山靛 Mercurialis leiocarpa, 箬叶竹 Indocalamus longiauritus, 香附子 Cyperus rotundus, 栗褐薹草Carex brunnea, 堇菜 Viola verecunda, 披针薹草 Carex lanceolata, 天目山薹草 Carex tianmushanica 等。
2、 研究方法
2.1 样地设置与调查
2012 年 10 月在龙王山自然保护地区, 选择地势较为平坦 、 植被类型典型的地段设置 1 个面积为100 m × 100 m 的固定典型样地。 用相邻网格调查方法, 把样地划分为 400 个 5 m×5 m 的调查单元。 在每个调查单元内, 对所有胸径在 5 cm 以上的活立木进行定位, 并进行每木检测, 测量并用号码牌为其编号, 逐株记录树种的名称、 胸径、 树高、 冠幅等测树因子。 同时记录群落中的生态要素。
2.2 物种重要值计算
重要值是一个比较客观的数值, 能较充分地显示出不同植物种群在群落中的地位和作用。 本研究采用的重要值计算公式为: 树种重要值=(相对多度+相对频度+相对显著度)/3。
2.3 数据分析
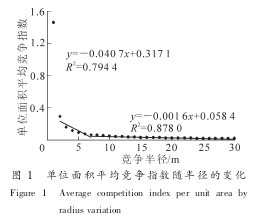
单木竞争指数的模型较多, 但以 Hegyi 的模型最常用, 预测效果最好。 本研究采用该模型计算竞争指数(competition intensity, CI)。 公式如下:
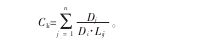
其中: CIi为第 i 株对象木的竞争指数; Dj为竞争木胸径; Di为对象木胸径; Lij为对象木和竞争木之间的距离; n 为竞争木的株数; j 为竞争木数量。
各优势种群的竞争指数用各优势树种林木点竞争指数之和表示, 公式如下:

其中: CI为某优势种群的竞争指数, 其值越大, 竞争越激烈; N 为对象木的株数。
在野外样地数据采集过程中, 发现有大量的无性分株现象。 本研究在数据处理时对无性系分株胸径大小的计算采用如下方法: 将同一基株上的各无性系分株按照胸高断面积求和, 然后再换算为一株胸高断面积与之相等的植株进行统计。
2.4 竞争影响范围
要描述林木之间的竞争, 首先要确定对象木周围竞争木的数量或竞争范围。 只有了解真正的、 具明显生态效应的竞争木的作用范围, 才能保证研究结果的准确性和可靠性。 在与位置相关的单木模型中,国内外的研究采用的样圆半径有 3.05, 6.00, 8.00 m 等, 这种方法易于计算, 但没有考虑处在半径边缘的林木因生长而成为新的竞争木的可能性。 也有许多研究采用 Voronoi 图来确定对象木的竞争木。
当竞争木距离对象木越远时, 生成的 Voronoi 图边数就越多, 故选择的竞争木的数量就越多, 这与事实不符。 对象木受的竞争压力来自其周围一定范围的个体, 随着距离的增加, 个体对它的竞争会减弱或消失。 所以, 本研究采用逐步扩大样圆半径, 以找出合适的竞争范围。 以 1 m 为样圆半径增量,分别计算 1~30 m 间 30 个样圆半径中对象木受到的单位面积平均竞争强度, 绘制出影响范围与单位面积平均竞争强度变化的散点图。 对不同影响范围进行分段拟合, 绘制线性回归图。 由图 1 可以看出, 在半径7 m 处有 1 个拐点, 竞争强度变化已经趋于稳定。
同时, 在样地调查中, 样地林窗半径约 6~7 m, 而且最大的树冠半径也没有超过 8 m, 故本研究采用的样圆半径是 7 m。
2.5 边缘校正
当样地内靠近边界的林木做为对象木时, 样地界外林木对它也产生影响, 这时, 需要进行样地边缘校正。
分别在原样地的上、 下、 左、 右、 左上、 左下、 右上、右下 8 个方向平移原样地, 形成由 9 个相同样地组成的大样地。 这样每个对象木都有 1 个相对完整的结构单元, 减少理论值与实际值之间的偏差, 消除边缘影响。
3、 结果与分析
3.1 优势树种组成及径级结构
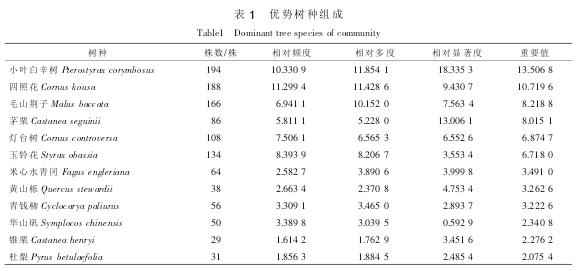
对样地调查数据进行统计, 结果显示样地胸径 5 cm 以上林木株数有 1 644 株。 计算各树种的重要值, 将各树种重要值由高到低排列, 重要值在 2%以上的有 12 种(表 1), 其中重要值大于 10%以上有 2种, 属于森林群落种的优势树种。 优势树种的株数和重要值分别占样地的 69.91%和 70.77%, 在群落中占明显优势, 决定着森林群落的结构和演替趋势。
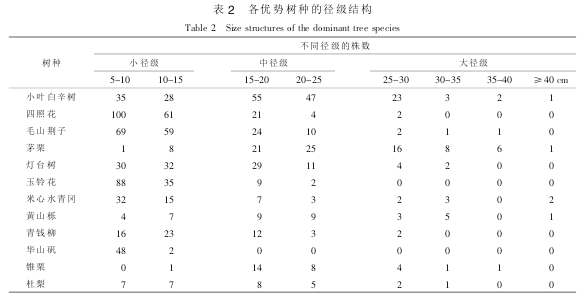
采用径级结构分析各优势种群的结构和动态。 按照林木的径级, 可以把林木分成 3 个级别: ①小径级组: 径阶 5~10 cm 和 10~15 cm。 ②中径级组: 径阶 15~20 cm 和 20~25 cm。 ③大径级组: 径阶≥25 cm。 从表 2 可以看出: 除了茅栗外其他各优势树种的株数均是随着径阶的增大而逐渐减小, 而茅栗株数随径级先增大后减小。 整体上, 各优势树种的中、 小径级林木居多, 其中, 小叶白辛树和四照花种群的中、 小径级林木相对其他优势树种较多。 各种群内更新资源充足。 这也说明群落总体上呈现稳定发展趋势。
3.2 优势树种种内竞争
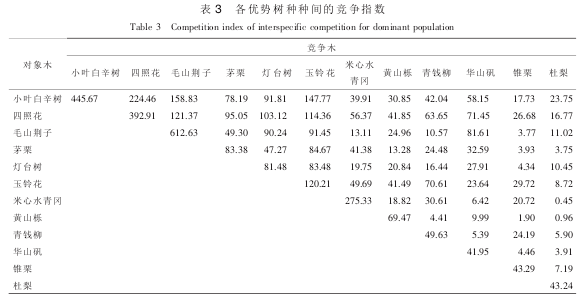
种内竞争是反映植物种内相互影响的一个重要特征。 样地中各优势树种的种内竞争关系如表 3。 各优势树种种内竞争强度有较大差别, 其大小顺序为: 毛山荆子>小叶白辛树>四照花>米心水青冈>玉铃花>茅栗>灯台树>黄山栎>青钱柳>锥栗>杜梨>华山矾。 根据各优势树种种内竞争强度的大小,可以把种内竞争强度分为 3 个等级: ①强度竞争(竞争指数>500), 优势树种只有毛山荆子。 ②中度竞争(200<竞争指数≤500), 优势树种有小叶白辛树、 四照花和米心水青冈。 ③弱度竞争(竞争指数≤200),优势树种有玉铃花、 茅栗、 灯台树、 黄山栎、 青钱柳、 锥栗、 杜梨和华山矾。 在样地调查中, 发现毛山荆子、 小叶白辛树、 四照花、 米心水青冈呈集群分布格局, 且毛山荆子、 小叶白辛树、 四照花、 米心水青冈种群的中、 小径级树占绝对优势。 中、 小径级树种正处在生长发育旺期, 此时, 环境的生长空间和营养条件均已不能满足植物个体的生长发育需要, 必然会向周围的树种发生争夺, 这样势必会引起周围大径阶树种对它产生较强的竞争, 故毛山荆子、 小叶白辛树、 四照花、 米心水青冈种内竞争强烈。 这也说明毛山荆子、 小叶白辛树、 四照花、 米心水青冈群落内将会产生自疏现象, 种内树木自然稀疏而加大距离, 使个体间对生长空间和营养条件的竞争性利用逐渐减弱, 而使林木逐渐趋于均匀化分布。
3.3 优势树种种间竞争
植物在生长过程中, 不仅与同种个体间发生竞争, 也与周围其他不同种植物不断地争夺水分、 光照、 空间及营养元素等, 因而产生了种间竞争。 种间竞争决定于该种的生态习性和生态幅度, 以及它们在生态系统中的生态位。 也就是说生态位接近的物种, 它们之间的竞争也就越激烈。 各优势树种的竞争强度不仅受竞争木的胸径和数量的影响, 也因竞争木种类不同而不同。
研究树种种间竞争时, 忽略竞争木和对象木的区别, 方法如: A 树种和 B 树种的竞争指数为对象木A 与竞争木 B 的竞争指数和对象木 B 与竞争木 A 的竞争指数平均值。 样地中各优势树种的种间竞争关系如表 3。 依据优势树种的种间竞争强度, 进行统计并排序, 划分为 4 个等级: ①种间竞争剧烈(147.77≤CI≤224.46)的有小叶白辛树和四照花、 小叶白辛树和毛山荆子、 小叶白辛树和玉铃花, 3 组。②种间竞争较强(63.65≤CI≤121.37)的有小叶白辛树和茅栗、 小叶白辛树和灯台树、 四照花和毛山荆子、 四照花和茅栗、 四照花和灯台树、 四照花和玉铃花、 四照花和青钱柳、 四照花和华山矾、 毛山荆子和灯台树、 毛山荆子和玉铃花、 毛山荆子和华山矾、 茅栗和玉铃花、 灯台树和玉铃花、 玉铃花和青钱柳, 14 组。 ③种间竞争中庸(11.02≤CI≤58.15)的有小叶白辛树和米心水青冈、 小叶白辛树和黄山栎、小叶白辛树和青钱柳、 小叶白辛树和华山矾、 小叶白辛树和锥栗、 小叶白辛树和杜梨、 四照花和米心水青冈、 四照花和黄山栎、 四照花和锥栗、 四照花和杜梨、 毛山荆子和米心水青冈、 毛山荆子和黄山栎、茅栗和灯台树、 茅栗和米心水青冈、 茅栗和黄山栎、 茅栗和青钱柳、 茅栗和华山矾等, 33 组。 ④种间竞争较弱(0.45≤CI≤10.57)的有毛山荆子和青钱柳、 毛山荆子和锥栗、 茅栗和锥栗、 茅栗和杜梨、 灯台树和锥栗、 灯台树和杜梨、 玉铃花和杜梨、 米心水青冈和华山矾、 米心水青冈和杜梨等, 13 组。
3.4 优势树种种内种间竞争比较
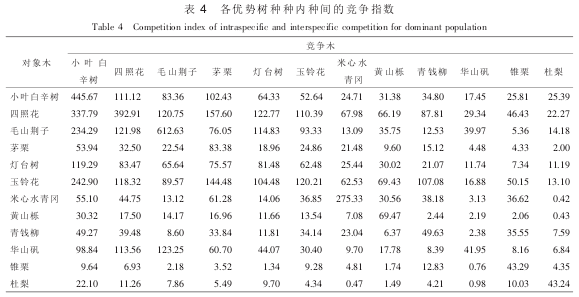
有学者采用 Hegyi 竞争模型分析常绿阔叶林种内和种间竞争强度时, 均发现常绿阔叶林优势种群的种内竞争比种间竞争激烈。 研究中以各优势种群分别为对象木时, 发现存在以下 3 种情况: ①种内竞争大于种间竞争的植物有: 小叶白辛树、 四照花、 毛山荆子、 茅栗、 米心水青冈、 黄山栎、 锥栗、杜梨。 ②种内竞争小于种间竞争的植物有: 灯台树、 玉铃花、 华山矾。 ③种内竞争与种间竞争相差不大的植物有: 青钱柳。 这也说明了落叶阔叶林种内和种间竞争状况比常绿阔叶林复杂。 一般来说, 同一个物种具有相同的生态位, 即它们之间对水分、 光照、 空间及营养元素有相同的需求, 故种内竞争相对种间竞争会激烈。 而灯台树、 玉铃花、 华山矾和青钱柳种群等的种内竞争小于或接近种间竞争, 这说明灯台树、 玉铃花、 华山矾和青钱柳种群等的生态位与小叶白辛树、 四照花较接近, 所以受到较强的种间竞争压力。 群落中种内竞争激烈的优势树种与其他优势树种的种间竞争也相对激烈, 如小叶白辛树、 四照花和毛山荆子。 而群落中种内竞争比较弱的优势树种与其他优势树种的种间竞争也相对较弱, 如黄山栎、 锥栗、 杜梨和青钱柳。 这与汤孟平等在天目山常绿阔叶林优势树种种间竞争分析中得出的结论是一致的。 通过表 4, 也容易看出小叶白辛树和四照花是其他优势树种最大的竞争者, 这主要是小叶白辛树和四照花作为群落中绝对优势的树种, 具有较多的数量且其生态位与其他物种较接近。
4、 结论与讨论
对龙王山落叶阔叶林优势树种种内种间竞争状况的研究结果表明: 研究的落叶阔叶林中优势树种有12 种, 包括小叶白辛树、 四照花、 毛山荆子、 茅栗、 灯台树、 玉铃花、 米心水青冈、 黄山栎、 青钱柳、华山矾、 锥栗、 杜梨等。 各优势树种的中、 小径级林木居多, 更新资源充足, 群落总体上呈现稳定发展趋势。 各优势树种种内竞争强度不同, 其中毛山荆子、 小叶白辛树、 四照花、 米心水青冈种群等的种内竞争比其他优势种要强烈, 属于中、 强度竞争, 这从侧面也反映出毛山荆子、 小叶白辛树、 四照花、 米心水青冈种群等呈现出一定的聚集分布状态。 依据优势树种的种间竞争强度, 划分为 4 个等级, 可为人工混交林树种选择提供参考。 灯台树、 玉铃花、 华山矾和青钱柳种群等的生态位与小叶白辛树、 四照花较接近, 故受到较强的种间竞争压力, 所以灯台树、 玉铃花、 华山矾和青钱柳种群等的种内竞争小于或接近种间竞争。 同时, 小叶白辛树和四照花等也是其他优势树种最大的竞争者。
Hegyi 竞争模型主要应用于纯林中, 它包括了胸径、 距离及竞争木数量等指标, 反映了林木个体对环境利用的差异, 是研究林木个体间竞争的适合量度。 本研究的结果表明: 将 Hegyi 竞争模型应用于龙王山落叶阔叶林优势树种的竞争, 能够较好地描述和揭示龙王山落叶阔叶林优势树种的竞争格局, 对推断和预测林分竞争状态, 为龙王山落叶阔叶林的管理与保护也具有一定的理论意义。 但是树木间的竞争能力受指数大小、 生长速度、 发育阶段等多种生物的和非生物因素的制约。 而 Hegyi 竞争模型在很大程度上代表林木间的地上部分竞争, 这也是它的不足之处。 所以, 要更准确地反映落叶阔叶林复杂的种内种间竞争关系, 需要更深入的研究, 比如在模型中增加一些树冠、 树高或环境因子等。
参考文献:
[1] 沈琛琛, 雷相东, 王有福, 等. 金苍林场蒙古栎天然中龄林竞争关系研究 [J]. 林业科学研究 , 2012, 25(3):339 - 345.
[2] 康华靖 , 陈子林 , 刘鹏 , 等. 大盘山香果树 Emmenopterys henryi 种内及其与常见伴生种之间的竞争关系 [J].生态学报, 2008, 28(7): 3456 - 3463.
[3] WEINER J. Neighborhood interference amongst Pinus rigida individuals [J]. J Ecol, 1984, 72(1): 183 - 195.
[4] 张学龙, 马钰, 赵维俊, 等. 祁连山青海云杉种群种内竞争分析[J]. 干旱区研究, 2013, 30(2): 242 - 247.
[5] MASAKADO K. Exploitative competition and ecological effective abundance [J]. Ecol Mod, 1997, 94: 125 - 137.
[6] 昭日格, 李钢铁, 岳永杰, 等. 浑善达克沙地天然沙地榆种内竞争研究[J]. 中国沙漠, 2011, 31(2): 451 - 455.
[7] WEINER J. Asymmetric competition in plant populations [J]. Trends Ecol Evol, 1990, 5(11): 360 - 364.
[8] 王晓霞 , 张钦第 , 毕润成 , 等. 山西稀有濒危植物脱皮榆种内和种间竞争 [J]. 生态学杂志 , 2013, 32(7):1756 - 1761.





