
动物卵母细胞体外成熟 (IVM)、体外受精(IVF)、卵胞浆内单精子注射(ICSI)和克隆等技术是动物胚胎工程的主要研究内容之一。近年来,家畜卵母细胞的体外成熟培养技术得到进一步提高,不仅为家畜胚胎生物技术提供了大量卵源,同时也是研究卵母细胞发生和发育规律的有效手段,但仍存在卵母细胞的成熟率、卵裂率及其囊胚发育率低等问题。为解决这一难题,秦鹏春等进行了大量的研究工作,认为FSH和LH这两种激素对猪卵母细胞IVM和排卵起到至关重要的作用.
SMAD3是TGF-β (transforming growth factor β)的下游受体激活蛋白。TGF-β超基因家族是在卵泡的生长、发育、闭锁及卵细胞的成熟和甾体激素的产生等方面都有重要的作用.SMAD3是其下游信号传导分子,TGF-β首先与TGF-βI(ITGF-βreceptor typeII)结合,迅速磷酸化TGF-βI(TGF-βreceptor typeI),继而激活SMAD3使其磷酸化,并与SMAD4结合转移入核,调节靶基因的功能.有大量实验表明,TGF-β/Smads通路对卵泡的发育、颗粒细胞的增值有重要作用,其变化可影响内分泌激素的作用,从而影响卵巢的功能.
开发优良家畜的生殖潜能,直接为社会提供优质的生殖配子或胚胎,必将创造更大的经济和社会价值。因此,本实以猪卵做研究对象,研究Smad3基因在猪卵母细胞成熟过程中的表达特征,为阐明Smad3基因与卵母细胞体外成熟的关系奠定基础。
1 材料与方法
1.1 材料
猪卵巢来自青岛市即墨屠宰场屠杀的青年母猪。采集卵巢后将其放入盛有青霉素、链霉素的37℃生理盐水保温瓶中,2 h内送回实验室进行卵母细胞的收集。将分别培养到0、24、36 h和48 h的COCs进行透明质酸酶消化,去除卵母细胞周围的颗粒细胞,Hoechst染色后在倒置显微镜下观察各个不同时期的细胞形态。卵母细胞GV期时卵内有一个较大的核,位于近边缘处,核膜清晰。GVBD期卵母细胞生发泡破裂,核膜消失,染色质凝集成团块状。MⅠ期卵母细胞染色体形成,并有规律的排列在赤道板上。MⅡ期卵母细胞最显着的特征是排出第一极体。
卵母细胞周围的卵丘细胞在结构上与卵母细胞相联系,参与GSH的合成。已有研究证明,卵丘细胞对于牛、猪和仓鼠卵母细胞中GSH合成起重要作用。
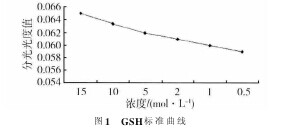
GSH在MⅡ期卵母细胞中的浓度大约是GV期卵母细胞中的2倍。在受精卵和胚胎早期发育阶段,GSH浓度迅速下降.GSH有还原型和氧化型(GSSG)2种形式。用分光光度计读取不同浓度谷胱甘肽的分光光度值,做成如图1的标准曲线。【图1】
1.2 方法
1.2.1 卵丘细胞-卵母细胞复合体的采集 对采集的卵巢用37℃的生理盐水清洗3~4次,用18号针头的20 mL的注射器,从直径3~6 mm卵泡的中抽吸卵丘细胞-卵母细胞复合体(COC)。为了保护卵母细胞,抽吸前在注射器中吸取少量预热的TCM199培养液。
抽完后将盛有COCs的50 mL离心管放置于38.5℃的水浴锅中静置以使COCs沉淀,每次静置15~20 min,弃上清液,用TCM199进行清洗2~3次,慢慢进行吹打,防止卵母细胞周围卵丘细胞脱落。将处理好的含有COCs的TCM199转入培养皿中,将培养皿置于体视显微镜下,捡出有3层以上卵丘细胞的COCs.
1.2.2 卵丘细胞-卵母细胞复合体的体外培养 把清洗干净的COCs用吸管转移至提前做好的培养滴中(6 cm培养皿,每个液滴150 μL,38℃,5% CO2,饱和湿度的培养箱)进行培养,其上覆盖矿物油。成熟培养液:TCM199培养液中添加L-半胱氨酸0.57 mmol/L、FSH 0.5 μg/mL、LH 0.5 μg/mL和 EGF 10 ng/mL.
培养0、24、36h和48h后,进行Hoechst染核(15min),在倒置显微镜下观察GV、GVBD、MⅠ和MⅡ4个不同时期卵母细胞细胞核的形态,统计各个时期的比率,并进行分类收集。
1.2.3 卵母细胞核质成熟的评定 细胞质成熟主要检测指标的是谷胱甘肽在4个不同时期(GV,GVBD、MⅠ和MⅡ)表达量的变化。用GSH检测试剂盒对GSH进行检测,按照Gasparrini等的方法,用紫外分光光度计测定猪卵母细胞内的GSH浓度。细胞核成熟检测指标主要是统计卵母细胞在培养48 h后排出第一极体的比率。
1.2.4 卵母细胞不同成熟时期 Smad3 基因 mRNA 的表达检测 将抽取的COCs,分别培养到0、24、36、48 h后消化分离卵母细胞和卵丘细胞,分别鉴定收集GV、GVBD、MⅠ和MⅡ的卵母细胞,利用单细胞RT-PCR技术检测Smad3基因的相应时期的表达情况。RT反应体系:5× RT Buffer,4 μL;Enzyme Mix,1 μL;047A primerMix,1 μL;RNase Free 双蒸水,4 μL. RT 反应条件:25℃,10 min;37℃,15 min;85℃,5 s;4℃,∞。实验采用β-actin 作为内参基因。扩增Smad3基因的引物序列如表 1. 反 应 体 系 为 cDNA,2 μL;SYBR Premix ExTaqTM(2×),12.5 μL;RNase-free water,9.5 μL;Forwardprimer,1 μL (5 μmol/L);Reverse primer,1 μL (5 μmol/L);共计20 μL.反应条件:95℃ 10 min;95℃ 15 s,60℃1 min,72℃ 10 s,循环40次。所用的数据均进行了中值标准化,平均数和标准差来源于3 个重复测量的结果,并且样品基因的相对表达量均进行了绘图。每个试验均重复3 次。计算公式为2^-(Cp目的基因-Cp内参基因)。【表1】

1.2.5 卵母细胞不同成熟时期 SMAD3 蛋白磷酸化水平检测 利用Western blot检测体外减数分裂成熟不同时期磷酸化Smad3的表达量,将COCs在体外培养0、24、36、48 h后,收集卵母细胞并Hoechst染核后,分别鉴定收集GV、GVBD、MⅠ和MⅡ的卵母细胞,用PIPA 裂解液和 SDS -PAGE 蛋白上样缓冲液处理,在冰上处理30 min,处理过程中每隔10 min,悬浮震荡1~2 min,裂解后,再加入4 μL SDS-PAGE 蛋白上样缓冲液,沸水处理5 min,未进行实验的样品可以置-20℃ 保存。实验中用到的一抗为Rabbit mab(按1∶200稀释),二抗为辣根过氧化物标记的山羊抗兔IgG(按1∶1000稀释)。
1.3 统计分析
卵母细胞GV、GVBD、MⅠ和MⅡ各时期的比例用ANOVA 方法分析。来自AVOVA 的重要结果再用Tukey 分析。Western Blot 结果图片扫描后,利用IPWIN 60进行灰度分析。40× 镜下进行各级卵母细胞计数,并将其比例利用SAS 软件进行方差分析。将Smad3基因的定量PCR 结果和GSH表达量测定结果利用SAS 软件进行方差分析.对所有的分析结果P<0.05为差异显着,P<0.01为差异极显着,每次实验至少重复3 次。
2 结 果
2.1 卵母细胞到达不同时期的比率 统计卵母细胞到达GV、GVBD、MⅠ和MⅡ4个不同时期的比率。如表2所示。【表2】
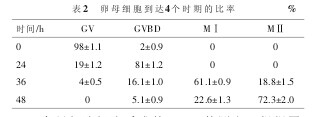
2.2 卵母细胞细胞质成熟-GSH的测定 根据图1标准曲线,计算出GV、GVBD、MⅠ和MⅡ4个不同时期GSH的浓度。在本实验中,培养的卵母细胞的谷胱甘肽水平随着培养时间的延长有所增高。说明在培养过程中卵母细胞细胞质成熟水平提高了。但是还不能达到体内GSH的水平,说明体外培养的环境还需要进一步提高。如图2、3所示,GVBD、MⅠ期与GV期相比差异显着(P<0.05)。MⅡ期时,GSH水平达到峰值,与GV期相比差异极显着(P<0.01)。【图2】
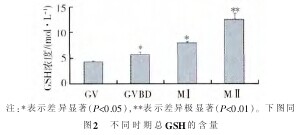
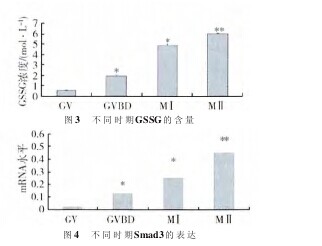
2.3 Smad3基因在卵母细胞不同成熟时期的表达情况 本实验旨在检测Smad3基因在卵母细胞发育过程中的表达情况,如图4所示,在卵母细胞成熟过程中的0、24、36 h和48 h,在倒置显微镜下捡取GV、GVBD、M Ⅰ 和 M Ⅱ 4 个不同时期的卵母细胞,通过单细胞RT-PCR检测,Smad3基因表达量随着卵母细胞的不同成熟阶段逐渐上升。GVBD和MⅠ期的mRNA水平与GV期相比,差异显着(P<0.05)。M Ⅱ 期 mRNA 水平达到峰值,与 GV 期相比,差异极显着(P<0.01)。【图3.图4】
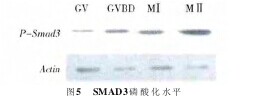
2.4 SMAD3蛋白在卵母细胞不同成熟时期的磷酸化水平 为进一步研究SMAD3信号通路在卵母细胞不同成熟时期的发育过程是否发挥作用,体外培养0、24、36 h 和 48 h 后,在倒置显微镜下捡取 GV、GVBD、MⅠ和MⅡ4个不同时期的卵母细胞,在蛋白水平上进行检测,结果如图5所示,随着培养时间的延长,P-SMAD3蛋白的表达水平是逐渐升高的,表明卵母细胞减数分裂成熟可能与Smad3基因的表达有关。【图5】
3 讨 论
3.1 影响
猪COCs体外发育的因素 影响猪COCs体外发育的因素众多,在实验过程中发现,保存卵巢生理盐水的温度是影响哺乳动物卵母细胞体外成熟的影响因素之一,对卵母细胞的成熟起了重要的作用。
有关采集卵巢所用生理盐水温度的研究很多,从20℃、25℃、30℃、31℃一直到37℃、39℃均有研究.刑风英等报道,在22℃、30℃、37℃、38.5℃、40℃下保存卵巢,随着保存温度不断升高,卵母细胞的体外成熟、卵裂率和囊胚率明显下降,37℃的生理盐水保存猪卵巢的效果最好。Prather等也曾报道了25℃和39℃的生理盐水温度对卵母细胞发育潜力有影响。
在取卵时,应先在水浴锅内预热生理盐水达到所需温度,本试验在37℃生理盐水中保存猪卵巢,2 h内将卵巢运回实验室。但是由于屠宰场到达实验室的路程较远,再加上气温等外界诸多条件的影响,卵巢在到达实验室时,温度大约降低4~5℃,因此,取卵时生理盐水的温度要根据天气变化做适当的调整,预防温度过低,影响卵母细胞体外成熟效果。激素对猪卵母细胞的体外成熟起着关键的作用。在体内环境中,卵母细胞在内源激素的作用下逐渐生长、发育、成熟,最后在LH峰的作用下排出体外,FSH与卵丘的扩张和卵母细胞质成熟有关。激素在猪卵母细胞体外成熟的影响中,发现猪在无激素的存在时也趋于发生减数分裂恢复,但培养液中加入FSH和LH,会明显加速减数分裂,而且LH可促进细胞质成熟.
卵母细胞和颗粒细胞共培养的条件下,细胞有很好的生长发育情况,在裸卵的情况下,卵母细胞的生长出现停滞不前的状态。结果说明,颗粒细胞对卵母细胞的发育有至关重要的作用。它可以提供卵母细胞的发育所需要的各种营养物质,所以颗粒细胞在卵泡发育的早期起着支持细胞的作用。在体外成熟过程中,卵丘细胞和卵母细胞存在着一定的相互作用,两者之间有间隙连接,可以相互传递营养物质和信息,从而进一步促进卵母细胞的成熟。所以说,抽取完整的健康的卵丘细胞对卵母细胞体外成熟有至关重要的作用。有研究发现,在猪的卵母细胞体外成熟过程中,在不存在卵丘细胞的情况下,向培养液中添加一定量的谷氨酰胺,即使能够促进GSH的含量,但是能够阻滞胚胎发育。因此有学者认为,卵丘细胞的扩散可以作为卵母细胞体外成熟的判断标准之一。
3.2 卵母细胞成熟中 GSH 和半胱氨酸的作用
卵子的成熟包括核的成熟和细胞质的成熟。如果细胞质成熟不充分,那么卵胞质中就会缺少使鱼精蛋白二硫键分解和染色质解聚的因子,包括谷胱甘肽(GSH)、核质因子(nucleoplasmin)和细胞成熟促进因子(MPF)等,精子进入后,精核解聚受阻造成受精失败。因为核成熟进程影响细胞质成熟,而细胞质成熟对细胞核成熟又具有促进作用。体外核成熟的完成并不能保证细胞质的正常成熟,细胞核和细胞质的成熟具有协调性。所以检测卵母细胞中GSH的含量代表了细胞质的成熟状态。GSH在哺乳动物细胞中发挥着重要的生物学作用,参与卵母细胞的成熟.半胱氨酸是GSH的前体物质,在牛卵母细胞IVM时,疏基复合物能增加细胞内的GSH的浓度,并保持GSH高浓度状态,从而提高胚胎的发育率.
在猪卵母细胞成熟过程中,培养24 h和36 h时,卵母细胞内GSH的浓度差异是不显着,而在48 h时,相对于GV期,GSH的浓度显着增高,当卵母细胞进入MⅡ期时,GSH浓度达到最高峰。有研究证实,高浓度的半胱氨酸在成熟培养全过程中能显着影响囊胚的发育,用含半胱氨酸的培养液培养猪的卵母细胞,可提高猪卵母细胞的成熟率.细胞内GSH的浓度较低可能是卵母细胞发育能力较低的原因。确切地说,GSH是卵母细胞活性和发育能力的生化标志物。
3.3 SMAD3 对卵母细胞发育的影响
SMAD3 在发情前期、发情期、发情后期以及乏情期的大鼠卵巢均有表达,定位于原始和初级卵泡的卵母细胞,腔前卵泡和小有腔卵泡期定位于颗粒细胞,大有腔卵泡中表达变少。Smad3在TGF-β抑制T细胞的活化及增殖功能方面发挥重要作用.Wolfraim等研究急性T细胞性白血病患儿体内Smad3基因的表达情况,结果发现虽然Smad3 mRNA表达水平正常,但是患儿体内检测不到SMAD3蛋白质,研究证实,TGF-β对白血病患者淋巴细胞增殖的抑制作用是Smad3途径依赖的。Smad3属于受体激活的Smads,它主要转导TGF-β和活动素的信号。Smad3基因剔除研究部分阐明了Smad3基因在脊椎动物发育中的功能。有实验表明,在Smad3基因缺失的小鼠体内卵泡发育紊乱,窦前卵泡向窦卵泡的发育受阻,凋亡基因的表达改变,颗粒细胞凋亡增加,闭锁卵泡显着增加.还有研究显示,Smad3基因缺失的雌性小鼠缺乏正常的动情周期,不能正常排卵;体外培养Smad3基因缺失的小鼠窦前期卵泡对FSH的刺激反应极为有限,卵泡中颗粒细胞数量很少,卵巢和颗粒细胞中CyclinD2等基因的表达明显减少,FSH不能刺激其卵泡的早期发育。另有研究发现,表皮生长因子促进卵巢颗粒细胞的增值需要SMAD3的存在.研究结果表明,Smad3 基因mRNA 水平随着卵母细胞的发育表达量逐渐升高,在到达MⅡ期时,Smad3基因mRNA水平最高,与GV期mRNA水平相比差异极显着(P<0.01)。这表明卵母细胞体外发育过程中Smad3基因起着非常重要的作用。同时SMAD3蛋白的磷酸化水平也是随着培养时间的延长表达量是上升的趋势,但是本实验并未对SMAD3抑制剂等做相关的研究与探讨,只能说明卵母细胞不同成熟时期的发育可能是通过SMAD3通路发挥作用。
SMAD3在卵母细胞减数分裂成熟中的表达以及磷酸化变化是逐渐上升的,该基因在IVM过程中,促进细胞的减数分裂。为最终完善卵母细胞成熟的分子调控机制,实现对卵泡卵母细胞发育的精确调控,开发优良家畜的生殖潜能,促进优良家畜品种的培育奠定了基础。
参考文献:
[1] 周洪彬, 肖雄, 包磊, 等。 猪体外胚胎生产的研究进展[J]. 中国农业科学, 2005, 39(7): 1457-1463.
[2] 秦鹏春, 谭景和, 吴光明, 等。 猪卵母细胞体外成熟和受精的研究[J]. 中国农业科学, 1995, 28(3): 58-66.
[3] Evin J A, Yan C, Matzuk M M. Oocyte-expressed TGF-βsuperfamily members in female fertility [J]. Mol Cell Endocrinol,2000, 159(1): 1-5.
[4] Drummond A E, Dyson M, Le M T, et al. Ovarian folliclepopulations of the rat express TGF-β signaling pathways[J]. MolCell Endocrinol, 2003, 202(12): 53-57.
[5] Itoh S, Itoh F, Goumans M J, et al. Signaling of transforminggrowth factor-beta family members through Smad proteins[J]. EurJ Biochem, 2000, 267(24): 6954-6967.
[6] 卫晓红, 祁丽花, 徐健, 等。 Smad2/Smad3蛋白在大鼠卵巢颗粒 细 胞 中 的 表 达 及 FSH对 其 活 化 的 影 响 [J]. 解 剖 学 报 ,2007, 38(2): 205-208.
[7] Wang Z G, Yu S D, Xu Z R. Improvement in bovine embryoproduction in vitro by treatment with green tea polyphenolsduring in vitro mturation of oocyte [J]. Anim Reprod Sci, 2006,100(1-2): 22-31.





