
土壤微生物是土壤有机组分和生态系统中最活跃的部分,是维持土壤质量的重要组成部分,土壤微生物对土壤中的动植物残体和土壤有机质及其他有害化合物的分解、 生物化学循环和土壤结构的形成过程起调节作用。 土壤微生物指标能敏感地反应土壤质量的变化,是土壤质量评价体系中不可缺少的组成部分。 一般认为, 土壤微生物的数量在一定程度上反映了土壤的肥力状况。 土壤微生物群落及其数量受温度、水分、植物残体及植物生长的综合影响,并随着季节的变化而变化,与土壤结构、养分及植被也有密切关系。
石梅湾海岸青皮林是海南岛重要的天然海防林,为抵御台风、防治土地风沙化、保障当地人民生命财产安全等起着不可替代的作用。 同时,对人工海防林向天然海防林恢复与重建具有重要的示范作用。 为了揭示海岸青皮林能在贫瘠的滨海沙土定居, 并长期自然更新演替, 除了对青皮种群的生物学及生态学特性进行研究之外, 有必要对其土壤性质尤其是土壤微生物类群进行深入探究。 目前关于该区域土壤微生物方面的文献报道还很少, 仅见有关土壤固氮微生物的少量研究。 为此,我们选择与 C、N 养分转化有关的土壤微生物生理类群作为研究对象, 通过对石梅湾海岸青皮林土壤根际与非根际微生物的对比分析, 并与相应土壤的总 DNA 含量做比较,以揭示青皮林土壤微生物的分布特征并试图探索其原因, 以期为青皮林的保护和管理,海南海防林的更新改造和建设提供科学依据。
1 材料与方法
1.1 研究地概况
研究区位于海南岛东部万宁市沿海 (18°39′17′′~18°40′14′′N、110°14′53′′~110°16′46′′E),为国家重点珍稀植物保护区。 石梅湾海岸青皮林属于单优青皮(Vatica mangachapoi)林。 本区属热带海洋性季风气候,干湿季明显,年均降水 2 032 mm,5~10 月为雨季,11~4月为旱季,年均气温 24.5℃,月均气温分布为:7 月>4月>10 月>12 月。 研究区土壤为滨海砂土,母质为近代浅海沉积物,青皮林土壤 pH4.1~5.1。
1.2 试验方法
根据石梅湾景观分布的研究结果,选择受人为扰动较轻的青皮林为研究对象; 取样时间跨度为一个生长季, 分别为 2011 年 10 月、12 月和 2012 年 4 月和 7月;使用土壤采样器按 5 点采样法采取 0~10 cm(上层)和 10~20 cm(下层)的根际及非根际土壤土样;用 MPN法测定土壤氨化细菌、硝化细菌、亚硝化细菌、反硝化细菌及好气性纤维素分解细菌的数量。 土壤总 DNA提取用 Foregene Soil DNA Isolation Kit。 取 4 次采样实验数据的平均值分析不同林分不同土层各生理类群细菌的数量分布; 分别用同一样地各土层根际及非根际土壤同一次采样数据的均值分析不同林分各生理类群细菌数量的时间分布动态。
2 结果与分析
2.1 5类生理类群细菌的数量分布特征
在青皮林土壤中,5 类生理类群细菌数量差别巨大, 其中氨化细菌数量最多, 达 108~1010MPN/g 数量级,2 个土层分布差别不明显, 但 2 个土层中非根际土壤中氨化细菌的数量远高于根际土壤; 好气性自生固氮菌的数量次之,在 102~104MPN/g 数量级之间 ,且主要分布在上层土中, 好气性自生固氮菌的平均数量为37.84×103MPN/g, 下层土壤中该细菌的数量则仅为2.57×103MPN/g,2 个土层中根际多于非根际, 但两者数量差别不大; 反硝化细菌的数量明显减少,2 个土层分布差别不太明显, 在 325.34~350.01 MPN/g 之间,但上层土中非根际土壤中反硝化细菌的数量远高于根际土,而下层土中则相反;好气性纤维素分解细菌数量最少,且主要分布在上层土中,为 45.18 MPN/g 数量级,下层土中仅为 8.65 MPN/g 数量级, 但上层土中非根际土远高于根际土,下层土中根际与根外数量则相当;土壤中尚未检测到亚硝化细菌(表 1)。【表1】

2.2 5类生理类群细菌数量的时间分布动态变化
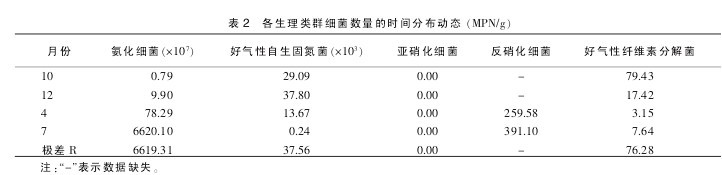
5 类生理类群细菌数量中,除亚硝化细菌在 4 次取样中均未检查到外, 其他 4 类细菌数量均随时间发生动态变化,但各自的变化规律明显不同。 氨化细菌的数量是 10 月最少, 仅为 0.79×107MPN/g, 经 12 月、4 月逐渐增多,到 7 月达到最大值,为 6620.10×107MPN/g,呈几何增长分布,极差值更是高达 6619.31×107MPN/g。
好气性自生固氮菌的数量变化为 12 月最多,为 37.80×103MPN/g,10 月次之,经 4 月 ,到 7 月降低为最小值 ,仅为 0.24×103MPN/g 数量级, 呈 A 型分布, 极差值为37.56×103MPN/g 数量级;反硝化细菌由于 10 月、12 月数据缺失,变化规律无法预测,但 7 月份高于 4 月份。
好气性纤维素分解细菌在 10 月数量最多, 为 79.43MPN/g,12 月次之,4 月最少, 为 3.15 MPN/g,7 月时略有回升,呈 U 型分布,极差值为 76.28×103MPN/g 数量级(表 2)。【表2】
2.3 土壤总 DNA 的提取及含量分析
提取 7 月 0~10 cm 土层土样的土壤总 DNA 并测定其含量,结果(图 1)显示,青皮林土壤的总 DNA 含量远高于木麻黄对照林 (该木麻黄海防林与海岸青皮林毗邻),其中青皮根际土壤的总 DNA 含量最高,每克土壤有 5.01 μg,其次为青皮林非根际土壤,为 4.37 μg/g,而木麻黄林根际土壤为 1.91 μg/g, 木麻黄林非根际土壤的总 DNA 含量最少,仅为 1.22 μg/g。【图1.略】
3 结论与讨论
微生物的种类及数量的分布与土壤的养分、 有机质含量、通气性以及水分含量等有密切关系。 土壤中氨化细菌主要发挥有机氮的矿化作用, 使不能被植物利用的有机含氮化合物转化为可给态氮, 为植物及其它微生物的繁殖和活动创造营养条件,故 5 类生理类群细菌中氨化细菌数量最多; 好气性自生固氮菌的生长不需可给态氮, 且其数量与土壤营养状况有密切关系,在土壤氮缺乏时,固氮菌才发挥固氮功能,为植物及其他微生物的生长代谢提供氮素营养,所以其数量次之;反硝化细菌的数量明显减少,好气性纤维素分解细菌数量最少。 除氨化细菌外,其他 4 个类群的细菌主要分布在 0~10 cm 土层中,好气性自生固氮菌、好气性纤维素分解细菌及亚硝化细菌都是好气性细菌,因此主要分布在 0~10 cm 土层中, 但是反硝化细菌作为厌氧菌, 却多数分布于 0~10 cm 土层中的原因还需进一步研究。
从细菌数量的时间分布动态变化来看, 氨化细菌和好气性纤维素分解细菌数量受气温影响较为明显,气温高时, 氨化细菌数量多而好气性纤维素分解细菌数少,其原因可能是 7 月高温多雨,有利于调落物降解而产生大量的有机态氮, 因此矿化转化所需的氨化细菌数量自然就多些; 而高温高湿的环境条件有利于纤维素分解真菌的生长, 此时凋落物的分解者主要是各种真菌, 纤维素分解细菌的数量受到抑制, 当气温下降,空气湿度降低时,不利于真菌生长,好气性纤维素分解细菌此时发挥主要作用,其数量增多。 因为土壤自生固氮菌数量与土壤营养状况及植被覆盖度有密切关系,所有好气性自生固氮菌数量 12 月最多,可解释为此时海南的气温较低, 不利于其他氮循环细菌如氨化细菌的生长,而植物生长继续消耗土壤氮营养,造成了此时土壤氮缺乏, 所以大量的固氮菌就发挥了固氮功能, 为植物及其他微生物的生长代谢提供氮素营养。
土壤自生固氮菌数量 7 月最少, 其原因可能是 7 月正值高温多雨季节,有利于土壤中各种凋落物降解,储存于凋落物中的氮营养大量返还土壤, 抑制了土壤好气性自生固氮菌的生长繁殖,所以此时其数量最少。
青皮林土壤中尚未检测到亚硝化细菌, 但不能确定土壤中一定没有亚硝化细菌存在。 这是因为土壤中的亚硝化作用和硝化作用是一个连续的过程, 亚硝化作用产生的亚硝态氮会被土壤中的硝化细菌迅速转化为硝态氮;另外,用 MPN 法测定亚硝化细菌数量是以检测亚硝态氮的存在与否为其数量指标的, 如果土壤中的亚硝化细菌作用产生的亚硝态氮被完全转化为硝态氮,或者是反硝化细菌消耗掉亚硝态氮,就检测不到亚硝态氮的存在。
分析结果显示根际土壤总 DNA 含量明显高于非根际土壤,说明根际土壤微生物量比非根际土壤多,与文献的研究结果一致。 但本研究所测定的各生理类群细菌中,氨化细菌、反硝化细菌和好气性纤维素分解细菌数量在青皮林中主要分布于非根际土中, 仅好气性自生固氮菌数量则在根际土壤中略多。 土壤中纤维素主要来源于植物凋落物, 而凋落物主要存在于非根际土中, 这就决定了好气性纤维素分解细菌主要存在于非根际土中。 其他生理类群细菌与植物根系的关系暂不明确,需进一步研究。
参考文献:
[1] 黄昌勇. 土壤学[M].北京:中国农业出版社, 2010.
[2] 孙波,赵其国,张桃林,等. 土壤质量与持续环境 Ⅲ.土壤质量评价的生物学指标[J]. 土壤,1997(5):225-234.
[3] 刘宛秋,杨家林.重建森林对退化红壤微生物特性的影响[J].福建林学院学报,2003,23(l):65-69.
[4] 崔炳华,蒋勇,陈绍武,等.巨桉人工林根际可培养土壤微生物数量的季节变化[J].四川林业科技,2012,33(5):63-66.
[5] 郝清玉,刘淑菊. 石梅湾青皮林群落结构及天然更新特征[J].林业资源管理,2010(2):54-59.
[6] 王贵,赵骥民,郝清玉,等. 不同植被土壤好气性自生固氮菌的时空变化格局及其菌株的初步筛选[J].广东农业科学,2013(13):60-64.
[7] 王贵,赵骥民,郝清玉. 石梅湾青皮林自然保护区景观破碎化研究[J]. 广东农业科学,2012(11):171-174
[8] 汪永华,胡玉佳,翁应云. 海南岛青皮林自然保护区[J]. 植物杂志,2003(5):8.





