����ժ Ҫ����������ĸ�����һ��������ض�����������ز��ܶ�����绷������Ӱ�����̬���ٵ����������������е����Ƹ��ܾ�֮һ�����Ǽٵ�����������������¸�������Ȼȱ����Ϊϵͳ���ܽᣬ�����Ƕ�Ӱ����ĸ��ܵ���ػ�����ؼ�������δ�ж��ۡ���˱������ȸ����˼ٵ����������ԡ����༰�ּ���죬���������˼ٵ���������Ⱦ��Դ�����ӵ������ã��Ǵ�л���������л��֬��л�ȣ��Ƕȷ����˼ٵ���������ˮƽ�ϵ��¸�ԭ�����е���ED;�������ø��Ļ����ذ��⵰��ø��AprX����ͱ���ABCת�˵��Ļ�����֬��ølipA�Ļ����ڲ�ͬ�����ºͲ�ͬ�����б������IJ��죬�����ǵ��¼ٵ��������ܵĸ���ԭ��ͨ�����ĵ��ܽᣬ����Ϊ��ʾ��Ʒ�мٵ������ĸ��ܻ��Ʋ�Ϊ���Ƹ����ṩ���ۻ�����
�����ؼ��ʣ�����; ����; �ٵ�����; ����;
����Abstract����Spoilage of raw meat is an ecological phenomenon closely related to many specific microorganisms and spoilage is affected by many external environmental factors. Pseudomonas spp. are one of is the dominant spoilage microorganism in raw meat. However, the mechanism of Pseudomonas spp.in the spoilage of raw meat is still lacking a systematic summary, especially the regulation and expression of genes involved in the spoilage of meat has not been determined. Therefore, this paper firstly summarizes the characteristics, classification and interspecific differences of Pseudomonas spp., and then overviews its contamination sources. It also elaborates the spoilage mechanisms from the perspective of substrate utilization (including carbohydrate metabolism, amino acid metabolism, lipid metabolism, etc.), and especially involved in the genes related to meat spoilage. Among them, the different expression levels of genes regulating enzymes in the ED pathway, and AprX gene that regulating extracellular protease and genes encoding ABC transporter, and lipA that encoding lipase may be the cause of the spoilage of Pseudomonas spp. The information gathered here will reveal the mechanism of Pseudomonas spp. on the spoilage of meat and provide a theoretical basis for controlling meat spoilage.
����Keyword����meat; spoilage; Pseudomonas spp.; mechanism;
������ĸ��ܱ�����ָ������������������£��ر����������Ⱦ��ʹ��Ӫ���ɷֺй���״�����仯�������ܲ����������к����ʵĹ���[1]��ȫ��������ÿ���ܶ��ܾ��������������Ϊ�������[2]����ˣ��˽����и����������Դ�����༰Ӱ����������ֳ����绷�����أ���ȷ��Ч���Ƹ���������������������ҵ���ѧ���繲ͬ��Ŀ��֮һ[3]��
�����ٵ�����(Pseudomonas spp.)�����ฯ�ܹ���������Ҫ���ܾ�֮һ�����ּٵ�������Ϊ���е����Ƹ��ܾ����������ӫ��ٵ�����(P. fluorescens)��ݮʵ�ٵ�����(P. fragi)��¡�¼ٵ�����(P. lundensis)������ٵ�����(P. putida)�ȡ��������������ԣ����������������������ĸ��ܱ��ʣ��ٵ��������Բ�������ø����������ø��֬��ø�ȣ���Щø���ǵ���������ʵ�ԭ��Ŀǰ�ܶ��о�̽���˼ٵ��������¸����ƣ���ȡ��һ���о���չ����˶Լٵ���������������¸����ƽ���ϵͳ�ܽᣬ���ڿ�����ĸ��ܽ���ʮ�ֱ�Ҫ��
������ˣ����ĸ����˼ٵ����������Լ�����Ⱦ��Դ�����ӵ������ã��Ǵ�л���������л��֬��л�ȣ��Ƕȷ����˼ٵ���������ˮƽ�ϵ��¸�ԭ������Ϊ��ʾ��Ʒ�мٵ������ĸ��ܻ��Ʋ�Ϊ���Ƹ����ṩ���ۻ�����

����1����ĸ���
������ĸ��������������Դø�Ĺ�ͬ���������[4]������Ҫԭ����������������ʱ����ӳ�����ı���������ֳ�������ý�����Ӫ�����ʲ�����л���������¡������������õ�Ӫ��������Ҫ��Ϊ�����ࣺ̼ˮ��������ǽͽ�;���е����õ�����������ԭ�������ǵȼ���л�������������Ρ�������-6-���ᡢ��ͪ���Ρ�����ȣ��������ʼ������ᣨ�����ᡢ�����������Ȱ����ᣩ��֬����֬�����[5,7]�����еĸ������ォ̼ˮ�������������ǵ����ʷֽ�Ϊ�����л��ᣬͬʱ��������CO2�����壻�����ĵ���ø��ʹ���еĵ����ʱ�ˮ�����ɰ����������⡢�ӡ�����ء�����[6]��������֬��ø��ʹ֬�����ֽ�����֬���ᡢ���͡�ȩ��ͪ�Ȼ����Ҳ���о���ʾ��Ʒ�ĸ��ܲ�����ȫ���������Ⱥ�����ã������ڳ�ʼ��Ⱥ��ռ������λ���ٲ����ض��������Ⱥ��һ���Ϊ���Ƹ��ܾ��������¡����Ƹ��ܾ�Ⱥ�����ࡢ����Ӱ������ĸ��ܽ���[1,9]��
����2���ٵ�������
����2.1�� �ٵ�����������
�������еĸ��ܾ�����࣬��Դ�㷺����Ҫ�����ٵ���������ϣ���Ͼ����������˾����ȸ��������Ծ�������˾������������������˿�����ȸ��������Ծ���[7]�����мٵ������������ฯ�ܹ���������Ҫ���õ����Ƹ��ܾ�֮һ���þ�Ϊֱ������ĸ��������Ը˾������ϸ����������һЩ�ٵ�����Ҳ�����������Ρ��������λ���������������ڵ�ȱ������������[8]��������Ӫ�������ϵ�һ����������ھ�����˫ˮ��ø�������ò�ͬ���л�������������Ρ��������κ������ǵȣ���Ϊ̼Դ�͵�Դ���ṩ�������������ܴ�л����������ѣ�����H2S[7]����������¶�Ϊ25��-35�棬���д��������Ϊ��������ٵ�����������Ӧ������ϸ��Ĥ�ڵĸ�ˮƽ������֬�ʺ��������յ�Ӧ���Ķ��ֻ����й�[10]��
����2.2�� �ٵ������ּ����Ʒ��
�����ٵ������ڲ�ͬ�����������������ˮ�֡��������ֲ����������˻��������¾��м�����Ҿ����������ࡣ��1894�걻�������������������ɺ�ۼ�����̬ѧ��չ����DNA�����ɺ�DNA-DNA�ӽ�����Ӧ���ڼٵ���������[11]������������RNA-DNA�IJ�����������ٵ�������Ϊ5��rRNA��Ⱥ[12]�����мٵ�����rRNA��I ��(P. sensu stricto)�����ฯ��������أ���ͭ�̼ٵ�����(P. aeruginosa)������ӫ��ٵ������Ͳ��ַ�ӫ��ٵ�������ɣ���ʩ�ϼٵ�����(P. Stutzeri)������ٵ�����(P. Alcaligenes)�������ٵ�����(P. Pseudoalcaligenes)���Ŷ����ٵ�����(P. mendocina)��[13]���ܶ�ѧ���о�����������ĸ���������ص�ݮʵ�ٵ�����[14]��ӫ��ٵ�����[15]��¡�¼ٵ�����[16]������ٵ�����[17]�ȶ��������Ⱦ�⼰����Ʒ�����������港�ܣ�������ճҺ�Ͳ�����ζ������
����ݮʵ�ٵ�����������������������ȴ������Ƹ��ܾ����ڸ������еļ���ʽ���56.7%��79.0%֮��[1]�������������ر��Ƕ������һ����Ĺ����л������ζ��ζ�������Ļӷ��Ի��������ͪ�ࡢ���ࡢ1-ʮһϩ�����������[14]��Ercolini[14]�ȷ��ֲ�ͬ��Դ��ͬ���͵�ݮʵ�ٵ������������ж�����һ���ĸ������������䵰��ˮ����Լ�֬��������Žϴ���죬�ڶ�������ľ�����ļ�����ľ��군��ˮ������ǿ����������װ�Ļ��������£�ݮʵ�ٵ��������ֽ⼡��ͼ������������Ϊ���Ƹ��ܾ�Ⱥ����ӫ��ٵ������౻���[9]�������¡�¼ٵ��������ڸ������еļ����Ϊ40%[3]���ܲ���������ͪ������Լ����ࡢ���ᡢ1-ʮһϩ�ͻ�����ȡ��ٵ������������������������Ⱥ������������Ԫ�أ�����Դ����������������������ݮʵ�ٵ�������¡�¼ٵ�������ӫ��ٵ��������ڳ�����˲ʱ�����ţ���м��[18]������ٵ�����������Ϊ������������ĩ�ڸ���������ƾ�Ⱥ�����ڻ����������У����Ž�����������Ⱦ�����Ҫ����[19]��
����3���ٵ���������Ⱦ��Դ
����3.1�� ������
����������ˮ�ֺ����ϸߣ�Ӫ���ḻ���ڹ��������ӹ����������ܵ������������Ⱦ��������ĸ��ܱ�����Ҫ���ɳ�ʼ����ռ������λ�������Ⱥ�����ģ��������ʼ��������༰����������Ч������ĸ��ܽ��̡�����ѧ���о����ּٵ������������⡢ţ�⡢���⡢����ȳ�ʼ��Ⱥ�е����Ƹ��ܾ�Ⱥ֮һ[19,20,21,22]��
����Korsak[23]�о����������������봫ͳ�������IJ��첻��Ӱ������������������ض����������ģʽ�����䲻ͬ������ϰ��Ӱ����������������Ⱦ��������мٵ����������������ִ�����ʽ��δ�������Բ��졣����ͩ[24]���о���������ţ�������У��ٵ�������Ҫ��Դ�ڲ���̨�ͷָ�ߡ������[25]�о������⼦�ӹ����ļӹ��������⼦����ϸ����Ⱦ��DZ����Ҫ��Դ�������⼦�����ӹ��IJ�ͬ������������Ⱦ��Դ����ͬ���������ˮ�ʡ�������Ա���ӹ����ߡ�����ȣ��ٵ�������Ҫ��Դ��Ԥ��Ρ����ɵ�[19]�����о������ڲ������ߺͰ������������ٵ����������ѻ���[21]������������Ӧ����������Ⱦ�������������Ȧ���ָ�ߡ�������˵������������ָ����������Ҫ��������ȾԴ�����мٵ���������Ҫ��Դ�ڷָ�ߡ�������˵��֡�Drosinos ��[22]���������������ӹ��Σ�ȥ��̨������豸�����ʹ������̺�������������Ⱦӫ��ٵ������ĵط���������[26]�о������������żӹ�����Ľ��У��ٵ����������������������ļ���ϸ����Ⱦ�̶ȸ��ڶ�����
�������ϣ���Ȼ������ࡢ���������������̼���ǰ�������в�ͬ�����ٵ������ڸ������ھ��в�ͬ�̶ȵļ������������������������Ǽٵ���������Ҫ��Դ֮һ�������ָ������Ӧ�������ߡ����˵��ֵȻ�����Ʒ�Ķ�ʱ��ϴ�������������淶�������̡�������Ա�߶����Լ��ټٵ���������Ⱦ��
����3.2 ����ͬ���ػ���
�����ٵ����������������ϼ����ϸ�����������ڵ����������ص����м�����˴��ε�[27]�о�����ţ����0 ����������£��ٵ��������¸��Խ�ǿ�����Ƹ��ܾ������ڸ��ܺ���ռ��������λ����հ�װ�����£�����������������ȱ�����������Ƽٵ������������������в���ѧ�߷�����հ�װ�����¼ٵ�����������ǰ�ڼ����ڸ���������ǿ���ں����ܵ����ơ�Chen��[28]����հ�װ�������������̽����ţ��ĸ��ܱ仯�����ּٵ����������������ڼ�������������½������ơ�Blixt��[29]�õ��ӱǶ���������հ�װţ��ı��ʳ̶ȣ���������ʼ�ٵ��������ڵ�����£�����ţ�ⸯ�̶ܳȵ����ӣ��ٵ��������������ܵ����ơ�������װ������ɷֱ�����ͬ��Ҳ��Ӱ��ٵ�����������״��������Х����[30]���о����ּٵ������Ǹ���������װ(80%O2/20%CO2)��ȴţ������ƾ��֣�����������ʱ����ӳ���������������һ����̼������װ(0.4%CO/30%CO2/69.6%N2)�¼ٵ������������ܵ����Ƶ����м����Fei��[31]��������װ��75%O2/25%CO2��75%N2/25%CO2��������������ϼ���˼ٵ������������е����������ּٵ�����������������ʱ����ӳ����������ơ�
������ͬ�������װ���������������еļٵ���������״����ͬ����ͬ������������֮�䶼�����Բ��죬����������벻ͬ�ٵ��������¸�����ĵ��ؼ���������йء�
����4�� �ٵ������ĸ�������
����4.1�� ��������
���������У��ٵ���������Ӫ�����������ࣨ�����ǡ�D-�����ǵȣ��Ͱ�������,�����ӷ��Ժͷǻӷ��Դ�л����ࡢͪ�ࡢ���ࡢȩ���л��ᡢ����Ͱ��ȣ�,��������̱䡢��ճ����ζ���������У�������������D-�������Ǽٵ��������������õ�̼Դ����������[3]������������Ҳ�����о������������ฯ�ܱ�־��ָʾ���ʣ��������мٵ���������ͨ��2-ͪ-3-����-6-����������;��(Entner-Doudoroff��ED)��л;���ֽ������Dz��������������Թ�����������ֳ��
�������������������Ĵ�����ʱ���ٵ���������������Ϊ�µ�������Դ[33]��������ļٵ������ܷ��ڵ���ø�����е���ˮ����ԡ����ҵ�������Ӫ���������Ĵ���ʱ���ٵ��������������䵰��ˮ�������������ڲ���ʹ�ⷢ�������εĸ��ܣ������ⸯ�ܵĽ��̡�
�������⣬�ٵ����������ڵ�֬��ø����������Ʒ�д�֬���������ˮ��Ϊ���ͺ�֬���ᣬ�Ӷ����¸���[34]��
�����ٵ��������¸����������������л�ֽ����������������Ⱥ����õ�˳����һ�����������лˮƽ�IJ�����ܵ�����������ԡ��������ؼ�Ⱥ���Ӧ�ȵ�Ӱ���⣬�������ܵ��ٵ������¸�����ĵ��أ���ˣ������ε�̽�������ˮƽ�ı��������Ը���һ����ʾ��ͬ�ٵ����������ض��Ļ����³�Ϊ���Ƹ��ܾ���ԭ���Ĵӵ������ã��Ǵ�л���������л��֬��л�ȣ��Ƕȸ��������˼ٵ���������ˮƽ�ϵ��¸�ԭ����ͼ1��ʾ��
����ͼ1 �ٵ��������������и��ܻ���ʾ��ͼ
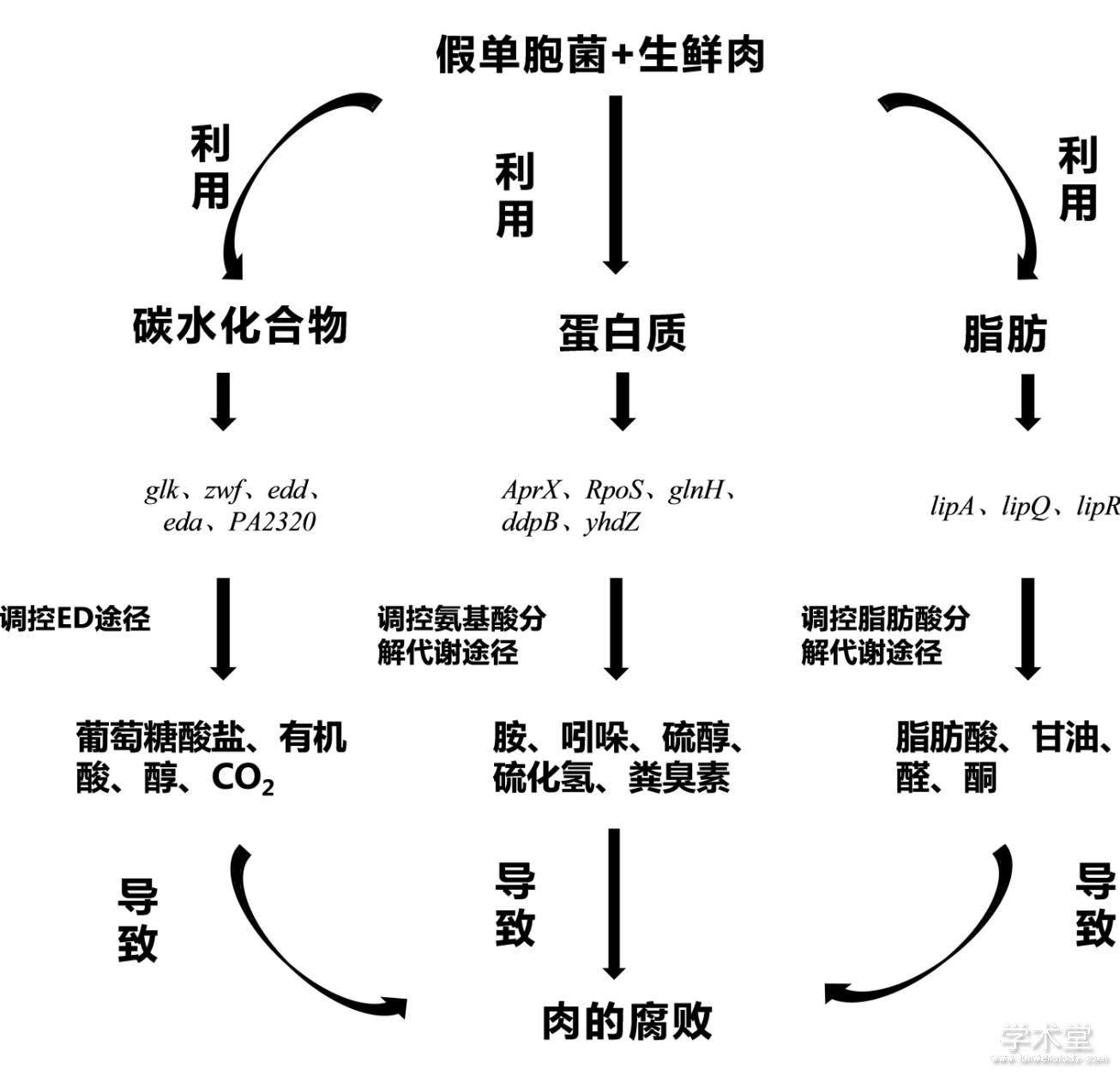
����Figure 1. Schematic diagram of the spoilage mechanism of Pseudomonas spp. in raw meat
����4.2���ٵ������븯����صĻ����о�
����4.2.1����̼ˮ�������л���
����һ����˵��ԭ��������ATP�ĺϳ���Ҫͨ���ǽͽ�(Embden-Meyerhof-Parnas��EMP)���������ữ����,����ѧ���о������ٵ���������ͨ��ED��л;������л�����ǣ��������������Ρ�Pablo I[32]���о����ֶ���ٵ�����������ȱ���ǽͽ�(EMP)ø6-�������-1-��ø(Pfk)���Ӷ�������ȫʹ��ED;�������⼺�ǡ�Castill��[35]���ֶ���ٵ������������Ƿֽ��лҲ��ͨ��3���μ�;��ͬʱ���У��ֱ��������Ǽ�ø;����2-ͪ��������ѭ���������������ữ��ͨ��������;��ʹ������ת����6-�������������Σ�����������Ĵ�лѭ�������б��������Ǽ�ø��glk����λ��edd����IJ������У�����������-6-��������ø��zwf1������eda�����γɲ����ӡ�Sudarsan��[36]���ֶ���ٵ�����KT2440ͨ��ED��EMP����������;����������(Tricarboxylic-Acid-Cycle��TCA)ѭ���������Ǵ�л��ԶԶ�����κ�һ���μ�;����GltR������������Ǽ�ø;����PtxS��LacIͨ������ͪ���������κ͵�������������;�������������ǵ����ã�ptxS�����lacI������ϵ����յ���������ȡ����oprB1��oprB2��PP1015��PP1018���ı����Լ��������������κ�ͪ����������ȡ�Ļ���gntP��kguT��kguK����ת������ʹ�����γ�����ED;���м���6-�������������εĻ���zwf1��zwf2��pgl�������ϵ����Ӷ�ʹ�ٵ������������ǵ��������ӡ���Abdelali[37]����Ϊͭ�̼ٵ�������ͨ��ED��л;������л�����ǣ����л�����ܵ������ض��ĵ���ϵͳ(PtxS��HexR��GtrS / GltR��GntR)�����������ǵĴ�л��ת�ˣ�����PtxS�������ӿ����������ı����Լ�������gadCBA��kguEKTD�ı�����߱������̼ˮ����������ʹ�л�ĵ����ʣ�HexR�������Ǵ�л�ϳ���ת����ص�glk��zwf��edd��eda����ı���[38]����ͭ�̼ٵ����������Ǵ�л��ػ�����Ŵ�ͼ����ʶ����ORF PA2320����Ƭ�Σ����Ӧ����Ϊת¼�������Ӽ����GntR[39]��GntR�������������β�����gntRKPZ�������Ǽ�ø(GntK)��������������ø(GntP)������������6-��������ø(GntZ)�ı��
����Mohareb��[33]���ֶ���ٵ������븯��������صĻ�����������������Ũ�����¶ȵĵ��ء��ڲ�ͬ�¶���PP4444��PP4030��PP3088����IJ����������������Ũ��Ӱ���������PP4444�����DNAת��ø�ı���������Ҫ��PP4030�������˳ʽ������֬����Ľ��⣬PP3088�����为����صĵ�������δ��֪���������ǵ����û���ֵ�ý�һ�����о���
����Wang��[40]ͨ��ݮʵ�ٵ������ڲ�ͬ��װ��ʽ�µ�ת¼�����������������װ�µ��Ӵ�����(nuoAB)����ı����ܵ������ƣ��ǽͽ�ͨ·�б�����������ȩøII���fbaA����Ƭ�ι�������Ӷ����ܵ��±�ͪ��Ļ��ۣ������˼ٵ������������ǵ����롣
��������Ʒ���ܳ��ڣ��ٵ������������е�������ͨ��ED;����EMP;������������;����TCAѭ��������ͨ·;�����������Ǵ�л���������漰�������Ǵ�л��صĻ���϶࣬Ŀǰ������ȷ���������Ǵ�л�����ٻ���ͬʱ�������ⸯ�ܵļٵ���������϶࣬ÿ�ּٵ������Ǵ�л�����ʡ�;��������ͬ�����ּٵ��������Ǵ�л;���Ƿ���ڵ����ⸯ�ܵĹ�ͬ�����д��ڽ�һ���о����ٵ������Ļ���Ƭ��ͨ������������Щ;���������Ǵ�л�Ĺ��̣���������������������Ϊ�ٵ�������������л�ṩ̼Դ������ʱ���㿪ʼ���ð�������Ϊ�µ�������Դ��
����4.2.2���백�����л���
���������о������������������£���������ĸ��ܳ��ڼٵ���������������Ϊ���Ƹ��ܾ���������������ʱ����ӳ����ٵ������ĸ������Ƹ�������[41]�����������ٵ������н�ǿ�ĵ���ˮ����ԣ��ܹ��ڱ���Ӫ����������������뵽����ڲ������������µ�Ӫ�����������������������л�йء�
����������ٵ������ܷ��ڵ���ø���羫����ø������������ø����������ø�ȣ����ٵ���������ع����з��ڵĵ���ø�Ǿ������ȶ��Եİ��⵰��ø������Ҫ������AprX���壬����һ����AprX�������ļ���п��������ø����[42]���ڶ���ӫ��ٵ��������У�AprX���嵰��ø�ѱ�ȷ��Ϊ����ʳƷ���ܵ�Ψһ����ø��Wang��[40]������������װ�£�ݮʵ�ٵ�����NMC25�漰����������ϳɵ�һϵ�л�����˿���ᡢ�Ӱ��ᡢ�����ᡢ�������ᡢ���װ��ᡢ�춬���ᡢ�����ᡢ����ȱ������ϸߣ�ͬʱҲӰ���˱���ABCת�˵��Ļ�����glnH��ddpB��yhdZ�ȣ��ı���Ӷ��������ฯ�ܡ��������ת¼��ͨ��RpoS�������ģ�RpoS���������RNA�ۺ�ø���[43]������ѧ��[44]��ΪCbrAB˫���ϵͳ��̼Դ��Դ�ļ��ְ�����(���鰱�ᡢ���������)ͬ��������ĵ���ϵͳ��
����������Ĵ�л��Ϊ���ӣ����������Ǻ�֬����Ĵ�л������ء�������ԣ�����AprX���嵰��ø�븯�ܵ�ȷ����[42]�����Դӵ���AprX����ø�Ļ������֣�̽���ٵ������백�����л��صĸ��ܻ������ذ�����ϳɵĻ���ͱ���ABCת�˵��Ļ���Ĺ��������ܹ�����ݮʵ�ٵ������ĸ��ܽ����Ѿ��õ�֤ʵ[40]�������ػ������������֣���ӫ��ٵ�����������ٵ������ж��ⸯ�ܵĵ������ÿ�����Ϊδ�����о�����
����4.2.3 ����֬�����л���
�����ٵ�������������Ʒ�͵�����Ʒ�з��ڰ���֬��ø���ֽ����õ����е�֬������֬���ᡢ���͡�ȩ��ͪ�����ʣ����ʳ�︯�ܲ��������˲��õ���ζ��֬��ø��һ����Ҫ������ø������α/β��ˮ��ø������[45]��������ٵ��������ܷ���֬��ø����֬��ø���������ƺ�;��������δ��������[46]��
�����ڼٵ��������У�ͨ����lipA�������֬��ø���γɣ�Conor��[47]����ӫ��ٵ�����B52��֬��ø(lipA)����ͼ��Խ�������ø(aprX)����λ��ͬһ���ٿ��ӵ����ˣ�������ܵ��¶��Լ���ѹ˫���EnvZ/OmpR������ϵͬԴ��ĵ��أ�Zha��[48]����Gac/Rsm�ź�ת��ϵͳ�Լٵ�������lipA����ı����е������ã�����Gac-RsmEϵͳ��Ҫ����ֱ�Ӽ���lipA���룬��Gac-RsmAϵͳ���Ӽ�����lipA�ķ��롣Krzeslak��[49]����֬��ø�ڼٵ������еı�����LipQR˫�����ϵ�ĵ��أ�����lipQ��lipR��������ı��뵰������NtrBC����ĵ������ӣ�ͨ�����ض������μ�����������(upstream activator sequence, UAS)���������֬��ø������˫��ֵ���ϵͳ����֬��ø�����ӵ�ת¼��
��������������˿��������(acylhomoserine lactones, acyl-HSL)��Ⱥ���Ӧ(Quorum Sensing, QS)ϵͳ��֬��ø����ı����Ӱ������[45,50]���ٵ�����ͨ��I�ͷ���;������ATP-��ϵ��ס���Ĥ�ںϵ�������Ĥ������ɣ���II�ͷ���;�������ǻ��������ӡ�ø�����ȹ��ɣ����ڰ���֬��ø��Rosenau��[51]�о�����֬��ø�ķ���;����ϸ����ǰ֬��ø��26���������N-ĩ���ź������йأ�ȱ���ź����е�ӫ��ٵ�����ͨ��I�ͷ���;������֬��ø��ӵ���ź����е�ͭ�̼ٵ�����֬��ø��ͨ��II��;�����ڵġ����������[46]��Ϊ�ٵ���������֬��ø�ķ���;�����Ƿ�ӵ�л���acyl-HSL��QSϵͳ�йء�
������������������֬��ølipA��������ܵ��������ؼ�����˫���ϵͳ�ĵ��أ�����֬��ø�ķ���;����������Ҳ��Ϊ���ӣ���ͬ�γ�;����֬��ø�书�ܱ����������Բ��컹��Ҫ��һ�����о���
����4.2.4 ��������Ĥ𤸽���
�������еĸ��ܾ�����𤸽��ʳƷ�Ӵ������ʳƷ�������γ�����Ĥ������ܵ���ʳƷ����������Ⱦ�����̱����ڲ�Ӱ���Ʒ�İ�ȫ�ԡ��ٵ��������¸�����������Ĥ���γɼ�𤸽���������ϵ[52]��Amador��[44]���ֶ���ٵ�����CbrAB˫���ϵͳ����CbrB��ȱ����Ӱ������Ĥ���γɡ�������[53]ȷ����fleQ��fliA��rpoN�Ƕ���ٵ������кϳɱ�ë������Ļ���ϸ�����˶���𤸽����лת����������ǵIJ�����DNA�͵����ʵķ��ڵȶ��ֹ��ܶ�����Ĥ���γɾ�����ҪӰ�죬������Щϸ�����ܵĻ���������Ĥ�����������ܵ��ܶ����صĵ��ء�Tribelli��[54]�о��������ڵ��������£�anr�������ϸ���ľۼ����˶���Ӱ������Ĥ�γɵĵ�һ�Ρ�ӫ��ٵ������е�adnA������ͭ�̼ٵ�����fleQ�����ͬԴ��������ϸ�����˶����ɱ�ë�鵼���ڸ�����Ʒ�����𤸽[55]����������������Ҫ�ٵ�������ݮʵ�ٵ�������ӫ��ٵ�����������ٵ�������������Ĥ�γ���صĻ����������ⸯ��֮��Ĺ�ϵ��Զδ̽����
����5 ������
������ĸ���������������ȫ����������ɵģ�Ҳ���ǵ�һ��������ã����ض����Ƹ��ܾ�Ⱥ�Ĺ�ͬ���õĽ�����ٵ�������Ϊ���������ع����е����Ƹ��ܾ������¸��Ļ���ʮ�ָ��ӣ��������ܵ����ֻ�������Ӱ�죬���һ��ܵ����Ǵ�л���������л��֬��л����ź�ϵͳ�ĵ��ڡ�
����Ŀǰ�Ը��ܾ��о������뵽����ˮƽ���ٵ��������еIJ�ͬ�����ڲ�ͬ�����������¸��������������ţ��������ٵ������ּ�ĸ��ܻ���ı�������йء�Ŀǰ������Ա�̽����ͬ�ٵ������ּ䡢������¸��������������о������٣���ˣ�δ�����о����ԴӸ��ܻ������ˮƽ�IJ�����̽���ٵ��������¸����ơ��������ͻ�������̽���ٵ����������¸���صĻ��������ڽ�ʾ��ĸ�����������ԭ�Ӷ��ҵ����õ����ƴ�ʩ����������ĸ��ܣ��ӳ���Ʒ�����ڡ�
���������
����[1] NYCHAS G J E, SKANDAMIS P N, TASSOU C C, et al. Meat spoilage during distribution[J].Meat Science, 2008, 78(1-2): 77-89. DOI:10.1016/j.meatsci.2007.06.020.
����[2] Food and Agriculture Organization of the United States. Save Food:global initiative on food loss and waste reduction[EB/OL]. (2016-12-04) [2017-01-10]. http://211.85.197.33:80/rwt/02/http/P75YPLUGMFYT655TMH/save-food/resources/keyfindings/infographics/meat/en/.
����[3]��һ��, ������, �����,��.����ţ���еĸ����������[J]. ʳƷ��ѧ, 2018, 39(13): 296-303. DOI:10.7506/spkx1002-6630-201813043.
����[4] ���F. ��ͬ�����¶ȶ����ʼ��������ƷƷ�ʵ�Ӱ���о�[D]. ����: ����ũ�ֿƼ���ѧ, 2015: 1-2.
����[5] ����. ��Դ����ϣ���Ͼ���������ѧ��̼Դ��л�о�[D]. �Ϻ����Ϻ������ѧ, 2017��3-5.
����[6]��ѩ, �̵�, ����, ��.������Դ����ø���о���չ[J]. ʳƷ�Ƽ�, 2019, 44(01): 37-41. DOI:10.13684/j.cnki.spkj.2019.01.006.
����[7]���·�, ����, �Ŵ���, ��.���������Ʒ�и���ϸ���о�[J]. ũ��Ʒ�����밲ȫ, 2018, No.093(03): 23-31. DOI:CNKI:SUN:NYZL.0.2018-03-005.
����[8] ANDREANI N A, FASOLATO L. Chapter 2 – pseudomonas, and related genera[J]. Microbiological Quality of Food, 2017:25-59. http://211.85.197.33:80/rwt/02/https/MSYXTLUQPJUB/10.1016/B978-0-08-100502-6.00005-4.
����[9] DOULGERAKI A I, ERCOLINI D, VILLANI F, et al. Spoilage microbiota associated to the storage of raw meat in different conditions[J]. International Journal of Food Microbiology, 2012, 157(2). DOI:10.1016/j.ijfoodmicro.2012.05.020.
����[10] MORENO R, ROJO F. Features of pseudomonads growing at low temperatures: another facet of their versatility[J]. Environmental Microbiology Reports, 2014, 6(5):417-426. DOI:10.1111/1758-2229.12150.
����[11] PEIX A, MARTHA-HELENA RAM?REZ-BAHENA, ENCARNA VEL?ZQUEZ. Historical evolution and current status of the taxonomy of genus pseudomonas[J]. Infection Genetics & Evolution, 2009, 9(6):1132-1147. DOI:10.1016/j.meegid.2009.08.001.
����[12] FRANZETTI L, SCARPELLINI M. Characterisation of pseudomonas spp. isolated from foods[J]. Annals of Microbiology, 2007, 57(1):39-47. DOI:10.1007/BF03175048.
����[13] ALVAROPEIX, MARTHA-HELENARAM?REZ-BAHENA, ENCARNAVEL?ZQUEZ. The current status on the taxonomy of pseudomonas revisited: An update[J]. Infection, Genetics and Evolution, 2018, 57: 106-116. http://211.85.197.33:80/rwt/02/https/MSYXTLUQPJUB/10.1016/j.meegid.2017.10.026
����[14] ERCOLINI D, CASABURI A, NASI A , et al. Different molecular types of pseudomonas fragi have the same overall behaviour as meat spoilers[J]. International Journal of Food Microbiology, 2010, 142(1-2):120-131. DOI:10.1016/j.ijfoodmicro.2010.06.012.
����[15] MELLOR G E, BENTLEY J A, DYKES G A. Evidence for a role of biosurfactants produced by pseudomonas fluorescens in the spoilage of fresh aerobically stored chicken meat[J]. Food Microbiology, 2011, 28(5):1101-1104 DOI:10.1016/j.fm.2011.02.00.
����[16] LIU Yongji, XIE Jing, ZHAO Lijun, et al. Biofilm formation characteristics of, pseudomonas lundensis, isolated from meat[J]. Journal of Food Science, 2015, 80(12):M2904-M2910. DOI:10.1111/1750-3841.13142.
����[17]������,�շ���,�ϻ�Ƽ,��.���������ж���ٵ������ķ��������[J].ʳƷ��ȫ�������ѧ��,2017,8(11):4436-4438.DOI��CNKI:SUN:SPAJ.0.2017-11-066 .
����[18] MORALES P, FERN?NDEZ-GARC?A, ESTRELLA, NU?EZ M. Volatile compounds produced in cheese by, pseudomonas, strains of dairy origin belonging to six different species[J]. Journal of Agricultural and Food Chemistry, 2005, 53(17):6835-6843. DOI:10.1021/jf050717b.
����[19] ����, �·���. һ�꽵��ƶ����Ķ���ٵ������ķ���������������[J]. ������ѧ, 2015(4): 814-816.
����[20] ����. ���ʼ�������������������Ⱦ�����������ʼ����о�[D]. �Ϸ�: �Ϸʹ�ҵ��ѧ, 2017:10-21.
����[21] ���ѻ�. �������Ӧ������������[D]. �ɶ�: ������ѧ, 2015:33-37.
����[22] DROSINOS E H. Microbial and physicochemical attributes of minced lamb: sources of contamination with pseudomonads[J]. Food Microbiology, 1995, 12(3):189-19. DOI:10.1016/S0740-0020(95)80097-2.
����[23] KORSAK N, TAMINIAU B, HUPPERTS C, et al. Assessment of bacterial superficial contamination in classical or ritually slaughtered cattle using metagenetics and microbiological analysis[J]. International Journal of Food Microbiology, 2017, 24(17): 79-86. DOI:10.1016/j.ijfoodmicro.2016.10.013.
����[24] ����ͩ. �½���ţ���ӹ������۹�������Ҫ������������ĵ����о�[D]. ��³ľ��: �½�ũҵ��ѧ, 2014: 8-20.
����[25] �����. ���ʼ����Ʒ������Ⱦ�������䱣�ʼ����о�[D]. �Ͼ�: �Ͼ�ũҵ��ѧ, 2009:15-23.
����[26]������.���ǵ�����ͬ���ڸ���������Դ����[J]. ����ũҵ��ѧ, 2015, 43(33): 112-114. DOI:10.3969/j.issn.0517-6611.2015.33.0395.
����[27] �˴���,��ΰΰ,�����.����ţ�������о�Ⱥ�ṹ�����ƾ��¸��Եķ���[J/OL].ʳƷ��ѧ:1-10[2019-04-24].http://211.85.197.33:80/rwt/02/http/NNYHGLUDN3WXTLUPMW4A/kcms/detail/11.2206.TS.20181214.1404.064.html.
����[28] CHEN Xue, ZHANG Yimin, YANG Xxiaoyin, et,al. Shelf-life and microbial community dynamics of super-chilled beef imported from Australia to China.[J]. Food research international (Ottawa, Ont.), 2019, 120. DOI:10.1016/j.foodres.2018.11.039.
����[29] BLIXT Y, BORCH E. Using an electronic nose for determining the spoilage of vacuum-packaged beef[J]. International Journal of Food Microbiology, 1999, 46(2):123-134. DOI:10.1016/s0168-1605(98)00192-5.
����[30] ��Х��. ������һ����̼������װ����ȴţ����ɫ�ȶ���Ӱ��Ļ���̽��[D]. ̩��: ɽ��ũҵ��ѧ, 2018:56-58.
����[31] ZHAO Fei, JING Hongpeng, WU Xinling, et al. Effect of modified atmosphere packaging in combination with ice-temperature on quality of mutton during storage[J]. Food Science, 2015, 45(9): 3772-3778. DOI:10.1021/ma300394u.
����[32] NIKEL P I, MAX CHAVARR?A, FUHRER T, et al. Pseudomonas putida KT2440 strain metabolizes glucose through a cycle formed by enzymes of the entner-doudoroff, embden-meyerhof-parnas, and pentose phosphate pathways[J]. Journal of Biological Chemistry, 2015, 290. DOI:10.1074/jbc.M115.687749.
����[33] MOHAREB F, IRIONDO M, DOULGERAKI A I, et al. Identification of meat spoilage gene biomarkers in pseudomonas putida using gene profiling[J]. Food Control, 2015, 57: 152-160. DOI:10.1016/j.foodcont.2015.04.007.
����[34] �����, ���ƾ�. ϸ��֬��ø���������ص��о���չ[J]. ����ѧ��, 2015, 55(11): 1378-1384. DOI:10.13343/j.cnki.wsxb.20150117.
����[35] DEL CASTILLO T, RAMOS J L, RODRIGUEZ-HERVA J J, et al. Convergent peripheral pathways catalyze initial glucose catabolism in pseudomonas putida: genomic and flux analysis[J]. Journal of Bacteriology, 2007, 189(14): 5142-5152. DOI:10.1128/JB.00203-07.
����[36] SUDARSAN S, DETHLEFSEN S, BLANK L M, et al. The functional structure of central carbon metabolism in pseudomonas putida KT2440[J]. Applied and Environmental Microbiology, 2014, 80(17): 5292-5303. DOI:10.1128/AEM.0164.
����[37] DADDAOUA A, CORRAL-LUGO A, RAMOS J, et al. Identification of GntR as regulator of the glucose metabolism in Pseudomonas aeruginosa[J]. Environmental Microbiology, 2017, 19(9): 3721–3733. DOI:10.1111/1462-2920.13871.
����[38] DADDAOUA A, KRELL T, RAMOS J L. Regulation of glucose metabolism in pseudomonas: the phosphorylative branch and entner-doudoroff enzymes are regulated by a repressor containing a sugar isomerase domain[J]. Journal of Biological Chemistry, 2009, 284(32): 21360-21368. DOI:10.1074/jbc.M109.014555.
����[39] JAIN, DEEPTI. Allosteric control of transcription in GntR family of transcription regulators: A structural overview[J]. IUBMB Life, 2015, 67(7): 556-563. DOI:10.1002/iub.1401.
����[40] WANG Guangyu, MA Fang, CHEN Xiaojing, et al. Transcriptome analysis of the global response of pseudomonas fragi NMC25 to modified atmosphere packaging stress[J]. Frontiers in Microbiology, 2018, 9(1277). DOI:10.3389/fmicb.2018.01277
����[41] ������, ��ٻٻ, ������, �ź���. ����̲���������о��嵰����������Ⱥ������Ĺ������о�[J/OL].ʳƷ��ѧ:1-10[2019-03-07].http://211.85.197.33:80/rwt/02/http/NNYHGLUDN3WXTLUPMW4A/kcms/detail/11.2206.TS.20181213.1429.050.htm.
����[42] ANDREANI N A, CARRARO L, FASOLATO L, et al. Characterisation of the thermostable protease AprX in strains of pseudomonas fluorescens and impact on the shelf-life of dairy products: preliminary results[J]. Italian Journal of Food Safety, 2016, 5(4): 6175. DOI:10.4081/ijfs.2016.6175.
����[43] REPOILA F, GOTTESMAN S. Signal transduction cascade for regulation of RpoS: temperature regulation of DsrA[J]. Journal of Bacteriology, 2001, 183(13): 4012-4023. DOI:10.1128/JB.183.13.4012-4023.2001.
����[44] AMADOR C I, IN?S CANOSA, GOVANTES F, et al. Lack of CbrB in pseudomonas putida affects not only amino acids metabolism but also different stress responses and biofilm development[J]. Environmental Microbiology, 2010, 12(6): 1748-1761. DOI:10.1111/j.1462-2920.2010.02254.x.
����[45] ROSENAU F, JAEGER K E. Bacterial lipases from pseudomonas: regulation of gene expression and mechanisms of secretion[J]. Biochimie, 2000, 82(11): 1023-1032. DOI:10.1016/S0300-9084(00)01182-2.
����[46] �����, �ű���, �ȫ,��. �ٵ�������֬��ø�ķ�������ѧ�о���չ[J]. �й����﹤����־, 2015, 35(9): 114-121. DOI:10.13523/j.cb.20150916.
����[47] MCCARTHY C N, WOODS R G, BEACHAM I R. Regulation of the aprX–lipA operon of pseudomonas fluorescens B52: differential regulation of the proximal and distal genes, encoding protease and lipase, by ompR–envZ[J]. FEMS Microbiology Letters, 2004, 241(2): 243-8. DOI:10.1016/j.femsle.2004.10.027.
����[48] ZHA D, XU L, ZHANG H, et al. The two-component GacS-GacA system activates lipA translation by RsmE but not RsmA in pseudomonas protegens Pf-5[J]. Applied and Environmental Microbiology, 2014, 80(21): 6627-6637. DOI:10.1128/aem.02184-14.
����[49] KRZESLAK J, GERRITSE G, MERKERK R V, et al. Lipase expression in pseudomonas alcaligenes is under the control of a two-component regulatory system[J]. Applied and Environmental Microbiology, 2008, 74(5): 1402-1411. DOI:10.1128/AEM.01632-07.
����[50] REIMMANN C, BEYELER M, LATIFI A, et al. The global activator GacA of pseudomonas aeruginosa PAO positively controls the production of the autoinducer N-butyryl-homoserine lactone and the formation of the virulence factors pyocyanin, cyanide, and lipase.[J]. Molecular Microbiology, 2010, 24(2): 309-319. DOI:10.1046/j.1365-2958.1997.3291701.x.
����[51] ROSENAU F, JAEGER K E. Bacterial lipases from pseudomonas: regulation of gene expression and mechanisms of secretion[J]. Biochimie, 2000, 82(11): 1023-1032. DOI:10.1016/S0300-9084(00)01182-2.
����[52] ����. ��Ϻ�ض����ܾ����ﱻĤ���γ����Լ�����֬�ĵĿ�������[D]. տ��: �㶫�����ѧ, 2013: 14-16.
����[53] ������. ����ٵ�����KT2442��ë�ؼ����ػ���Ծ�������ѧ���Ե�Ӱ��[D]. ����: ���ϴ�ѧ,2018: 8-27. DOI��CNKI:CDMD:2.1018.063656.
����[54] TRIBELLI P M, HAY A G, NANCY I L?PEZ. The global anaerobic regulator Anr, is involved in cell attachment and aggregation influencing the first stages of biofilm development in pseudomonas extremaustralis[J]. PLOS ONE, 2013, 8(10): e76685. DOI:10.1371/journal.pone.0076685.
����[55] WILLIAMSON K S, RICHARDS L A, PEREZOSORIO A C, et al. Heterogeneity in pseudomonas aeruginosa biofilms includes expression of ribosome hibernation factors in the antibiotic-tolerant subpopulation and hypoxia-induced stress response in the metabolically active population.[J]. Journal of Bacteriology, 2012, 194(8): 2062-73. DOI:10.1128/JB.00022-12.





