����ժ Ҫ������2006��Takahashi��Yamanaka���������յ����ܸ�ϸ��(induced pluripotent stem cells, iPSCs)���������ܸ�ϸ�����������ǰ��δ�еķ�չ״̬���ڼ�����ģ��ҩ����Լ�ϸ���Ʒ��ȸ����涼������Ҫ���ã��ٽ���ϸ������ѧ������ҽѧ��ѧ�Ƶķ�չ��Ŀǰ��iPSCs�����ѳ�Ϊ�о��������Ƶ���Ҫ���ߣ�����iPSCs����ɸѡ����ҩ�������з��У�ʹ��iPSCs����ϸ�����ٴ���������Ҳ����������iPSCs�����༭�����Լ�3D���������ϵ������о���չ�ٽ���iPSCs�ڼ����о��еĽ�һ��Ӧ�á����Ľ����˽������ر�̷����ĸ��£����������ϲ�������ϵͳ�����ϷDz�������ϵͳ�������ϲ�������ϵͳ�Լ������ϷDz�������ϵͳ�����ر�̷��������ף�ͬʱ������iPSCs�ڼ�����ģ�Լ��ٴ����Ƶȷ���������о���չ��Ϊ�ٽ�iPSCs������������о��ṩ�ο���
�����ؼ��ʣ���iPSCs; �ر��; ������ģ; ϸ���Ʒ�;
����Abstract����Since Takahashi and Yamanaka reported the generation of induced pluripotent stem cells(iPSCs) in 2006, the field of pluripotent stem cells has entered an unprecedented state of development. It plays an important role in disease modeling, drug discovery and cell therapy, and promotes the development of cell biology and regenerative medicine. At present, iPSC technology has become an important tool for studying of pathological mechanisms. New drugs screened by iPSC technology are being developed, and the number of clinical trials using i PSC-derived cells is gradually increasing. The latest research progress of iPSCs, combined with gene editing technology and 3D organoid methodology, promotes the further applications of iPSCs in disease research. In this review, we introduce the innovation of reprogramming methods in recent years, analyze the advantages and disadvantages of four reprogramming methods: integrated virus vector system,integrated non-viral vector system, non-integrated virus vector system and non-integrated non virus vector system. At the same time, we summarize the latest research progress on iPSCs in disease modeling and clinical treatment strategies, so as to provide a reference for further in-depth research in various fields of iPSCs.
����Keyword����iPSCs; reprogramming; disease modeling; cell therapy;
�������������Ϊ������ij�����ϸ���Ļ����鱻������������ϸ��״̬�����ָ���������ȫ�Ķ�����״̬[1]��Ȼ����1958��Gurdon��[2]������צ�(Xenopus laevis)������ϸ����ע�䵽ͬһ���ֵ�ȥ����ĸϸ���У��ɴ˲�����һֻ�������Ƶ���39���Wilmut��[3]���ú���ֲ���������˵�һ�����鶯���¡��——“����”����Щ��¡�о��������ֻ�ϸ����Ȼ�����ŶԻ��巢��������Ҫ��“�Ŵ�”���䡣1981�꣬Evans��[4]������̥��ϸ��(embryonic stem cells,ESCs)���Դ�С�����ߵ���ϸ�����л�ã����Thomson��[5]����������ϸ���ųɹ����������̥��ϸ��(human embryonic stem cells,h ESCs)��ESCs�Ƕ��ܵģ����Ǿ��������Ƶ����Ҹ���������ͬʱӵ��������ϸ�����ͷֻ���DZ�ܡ�1987�꣬Weintraub��[6]���ֵ�������Myo D�ı�����Խ�С�����άϸ��ת��Ϊ������ϸ������һ����֤��ÿ��ϸ�����������������ڻ�����Щ�����ڻ�����ά��ϸ�����Ե����á�2006�꣬Takahashi��Yamanaka[7]��24����ͬ�Ļ������֣�ͨ����ת¼������������ϸ������ת¼���ӣ������������4��ת¼����Oct4��Sox2��Klf4��c-Myc (OSKM)���Խ�С�����άϸ��ת��Ϊ����״̬���Ӷ��������յ����ܸ�ϸ��(induced pluripotent stem cells,i PSCs)������

����2007�꣬�ձ�������ѧTakahashi�о�С���������˹���Ǵ�ѧYu�о�С��ֱ𱨵��˴��˳���άϸ���в��������յ����ܸ�ϸ��(hi PSCs)�İ���[8,9]���Դˣ�Ѹ�ٷ�չ������hi PSCs����Ϊ������ģ��ҩ��ֺ�����ҽѧ��������һ�������˷ܵ���ʱ��������hi PSCs�ڼ�����ģ��������ڴ�ͳϸ��ɸѡ�����ƣ���hi PSCsģ������ҩЧ��DZ�ڶ��Ե�ҩ��ɸѡ���Խ��Խ���С�hi PSCs���ŵ����������Դ�����ࡢ��á�����չ���ܹ������������������ϸ�����ͣ�������h ESCs��ص��������⣬�Լ����п������û����ض���i PSCs�������Ի�ҩ����и��Ի����Ƶ�DZ��������3D���������Լ�����༭�������ر���CRISPR/Cas9�����ķ�չ��hi PSCs�����༲����ģ�ͻ��ڸ�ϸ�����ٴ����Ʒ���չʾ�����ǰ����������Ҫ�������ر�̷����ĸ����Լ�i PSCs�������Ŵ�������ģ��ϸ�����Ʒ�������·�չ���ܽ���Ŀǰi PSCs�������ٵ���ս��̽���˽����Щ����ķ�����
����1�� �ر�̷���
����1.1 ��OSKMת¼���ӵ�����
����Oct4��һ������ܸ�ϸ��(PSCs)�Ķ�����ά���йص�ͬԴ��ת¼���ӡ�Sox2�ڿ���Oct4�ı���������������Ҫ������[10]����Nanog��Oct4һ���˶����ԵĹؼ�ת¼���硣c-Myc��һ������ְ�֢������ص�ԭ����������ļȾɫ�����ε��ף��շ��㷺��ת¼�������c-Myc��һ���������ԣ��������о�����ת�����Ե�L-Myc����Խ��������[11]��Klf4��Ϊ�������������������ӣ��ǰ�Ѫ���������ӵ����ΰе㣬�ܼ���Sox2�ı���[12]������ת¼��������ϸ��������������н�ϵ�������Ҫ��DNA�������鵰�����κ�ATP������Ⱦɫ�����ܲ�����Ⱦɫ�ʽṹ�仯�ĵ���[13]����Stadtfeld��Hochedlinger[14]�ı�����ϸ���Ķ����Ա��յ�ʱ�ἤ������ת¼�����ڵ�һ��ת¼���У�c-Mycͨ��������H3K4me2��H3K4me3����ϸ�����������ϣ���־��Ⱦɫ�ʵ����ܡ��������ϸ����ػ������ij�Ĭ������Thy1��Snai1��Snai2��Zeb1��Zeb2�ȼ���ʻ���[13,15]���ڶ���ת¼����������Ⱦɫ�����ܵ�ϸ����OSKM�������ڶ�����ػ���(pluripotency-associated genes,PAG)����ǿ�Ӻ������ӣ���������ת¼�ͱ���ڴ�ת¼���У���ϸ�����ȸı�����̬����ǿ��ֳ�������������������Ƥ����ת��(mesenchymal-to-epithelial transition,MET)�������Cdh1��Epcam��Ocln����Ƥ������ϵ�����Ƥ��������״̬�Ľ���[16]���γ��˸����ES��ϸ���š���MET��һ������͵�Ч�Ĺ��̣���Ϊ�����յ������ϴ��ڼ����鵰�ף��������鵰������Ⱦɫ�ʹ���[13]��
����Klf4���������ζ���������Ҫ�Ľ�ɫ�����ȣ��ڵ�һ��Klf4��ϲ��������E-cadherin���ڵ���Ƥ�������Ʒֻ�����[17]����Σ��ڵڶ��μ�����Դ��Oct4��Sox2�ı���Ӷ�����ά�ֶ���״̬�����ҵ��ڻ���Klf4ͨ������ϸ���ķ�������ֳ���ֻ��͵����ȹ��̣��ڶ��ܸ�ϸ���з�����Ҫ�����á�Klf4��Oct4��Sox2֮�䳬ǿ������ü���һ����Nanog��Esrrb��Klf2��Sall4��ZFP42��ת¼���ӣ��Լ���Smad1��Stat3���ź�ͨ·��������[18]��
����1.2�� �ر������
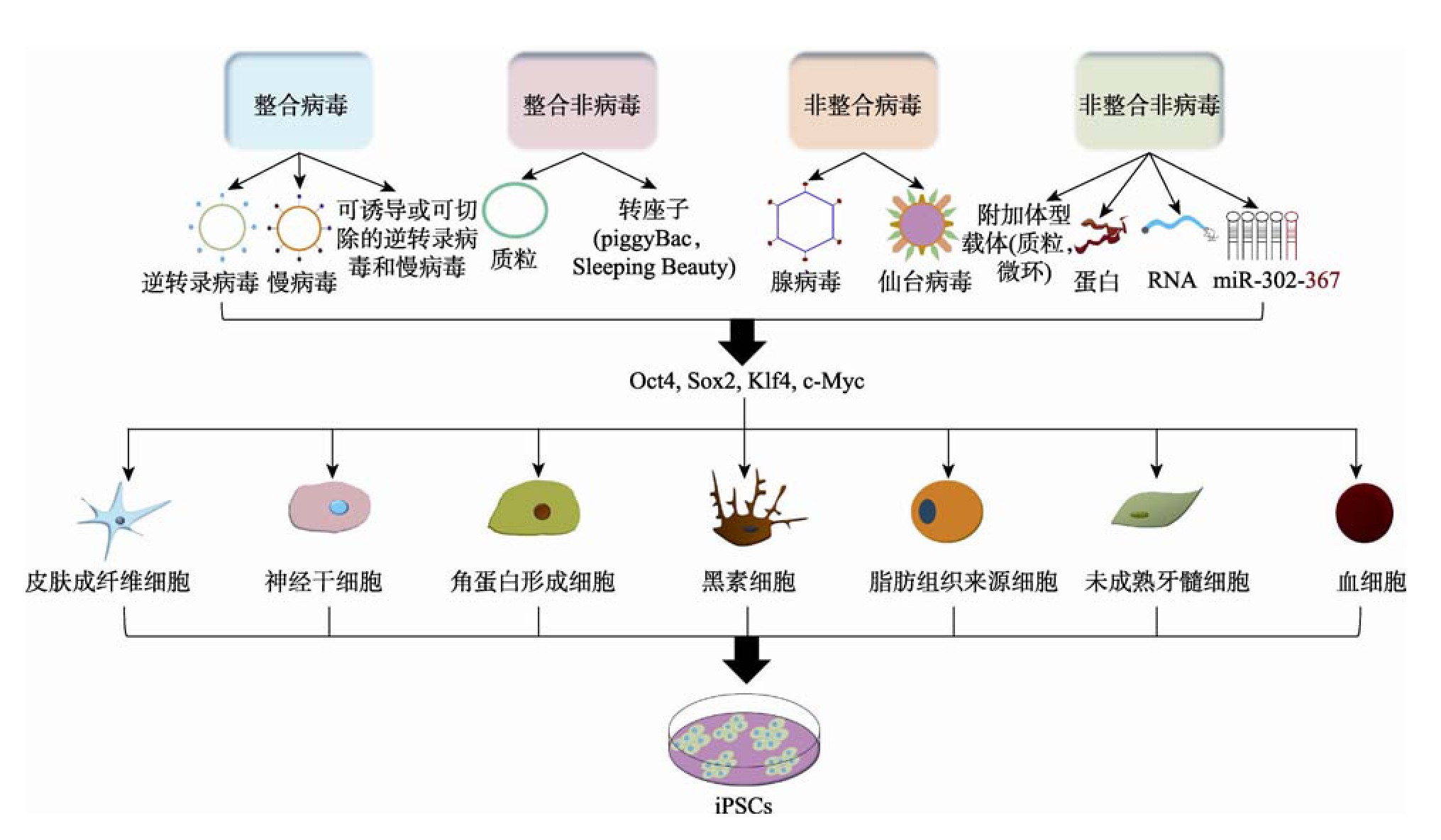
������Yamanaka�Ŀ������о�֮��½���м����о���ʹ�ò�ͬ�ķ���������i PSCs����������Ч����Ӧ�õİ�ȫ�Ժ�����������Щ�ر�̲��Դ��·�Ϊ���飺(1)ͨ���������ϻ������ת��ϵͳ��(2)���ò�����Dz��������������ر�̲��Խ����е��ر�̷�����Ҫ��Ϊ���֣����ϲ�������ת��ϵͳ�����ϷDz�������ת��ϵͳ�������ϲ�������ת��ϵͳ�Լ������ϷDz�������ת��ϵͳ��ͨ��4���ر�̷���������ϸ���ر�̳�i PSCs�Ĺ�����ͼ1��ʾ��ÿ�ַ�����������ȱ�㣬Ŀǰ��δ�ҵ�һ�������ƻ���DZ�ڲ�������ĵ�һ���ر��ת��ϵͳ��
����1.2.1�� ���ϲ�������ת��ϵͳ���ر��
����i PSCs�����ͨ����ת¼�������������ر�����Ӳ����ģ���ת¼���������ѱ��㷺��������������о��Ļ���ת������[19]�����ر�̹��̵ĺ��ڣ����ڱ����Ŵ�����[20]��ת�����ı��ﱻ��Ĭ[21]����ת¼��������ֻ�ṩ��ԴDNA���е�˲ʱ�����δ����ȫ�������������ص���Դ����ı��ͨ����ת¼�����������ɵ�i PSCs���������ܵ�������[22]�����⣬һЩ����ָ��������ת��������¼�����ڲ���i PSCs�еIJ������Ի�ı�ϸ���ķ������̣������ܵ���Ƕ�϶����������γ�[23,24]��
��������������(lentivirus,LV)����㷺�����Զ�����Ϊ����ת¼�������������Ч[24]�����LV�������ر��������ϸ�����͡�LV���ݷ��������������Ч���ر�̲���֮һ���ر��Ч��Ϊ0.1%��1%[14,25]������ѧ��������Ŭ��������һ;���İ�ȫ��[26]���������Ч���ر��LV����ȡ�õĽ�չ֮һ��������һ�ֶ�˳����LV������һ������ϵͳ��Я������Դ�Կ����߲�����2A“����”���������ӵ�һ���������������ĸ��ر������[23,27]����ϵͳ�����˲�����ת����ϸ�������ϵĿ�������������ת�����Ĭ�ķ��գ�����ת�����̣���������һ�µ��ر�����ӵĻ�ѧ������[28]�����⣬Ϊ��������Ч�ʳ�Ĭ��ת�����ټ����Ӱ�죬ͨ��������г�����(Cre/lox Pϵͳ)[29]�Ϳ��յ�ϵͳ(�Ļ���/ǿ��ù���յ�ϵͳ)[26,30]�����˳���Ӳ������壬���Ϻ��ת�������ͨ��˲ʱ����Cre������ϸ�����������Ƴ��������ֲ��Ե�ת��Ч�ʺܵ�[31]�����ҿ��ܵ���i PSCsͻ�䣬��ΪCre/lox Pϵͳ�����±�̺���ܻ�����lox P�ĺۼ�[32]��
����ͼ1 ����i PSCs�ĸ���ϸ����Դ���ر�̷���
����Fig.1 Various cell sources and reprogramming methods for the generation of i PSCs
����1.2.2 �����ϷDz�������ת��ϵͳ���ر��
��������Ŀǰ���ϲ���ת��ϵͳ�ľ����ԣ���ѧ����һֱ�ڻ����о������ر�̷�������Dz���ת��ϵͳ����Щ������������Ӧ�ø���ȫ����һ���ɹ��ķDz�����i PSCs���ɳ������̥����άϸ����������������תȾ�����ģ���һ����������c-Myc�����ڶ���������˳�����������OSKM�ر������[23]����Щ���ֱ���OSKM��˲ʱ�����������յ���ϸ���Ķ����ԣ��������ϵķ��պ��ر�̵�Ч�ʵ����ֳ�����Ҫ����[33]����ѧ�����������һ�����ϵ������Ի���ת�����壬��ת¼�������ϵ�����ṹ��lox Pλ�����Խ�����Ϸ��յ�����[28,29]��Ȼ�����̵�����Ƭ�ο��Դ������г���Ļ�����ϸ���У�����ܻ�Ӱ��ϸ������[33]�������ַ���������piggy Bac(PB)ת���ӵȿ��ƶ��Ļ���Ԫ����������Դ�����Ի���dz���Ч����PBת��ϵͳ����ת��Ч�ʸߡ�ɾ����ȷ���ŵ�[34]���˷���ͨ�����ݵ�תλ������Դ����±�̵�ϸ������ȫȥ����Ԫ���IJ���[35]�����ҵ��ǣ����������������Դ�Ե���PBת����Ԫ������ת�����г��������������ԵĻ�����ı�[36]��Ϊ�˷�PBת���ӵľ����ԣ�������������Sleeping Beauty(SB)ϵͳ��ʹ������Ƶ�ʵ���PBת���ӣ����������������û����SBԪ��[37]��Ȼ�����ֵ������µ����⣺�˷������ر��Ч�ʺܵͣ�ʹ�ÿ��г���Ԫ�������ܻᵼ���������ϵķ���[33]��
����1.2.3�� �����ϲ�������ת��ϵͳ���ر��
����Stadtfeld��[38]�����ٲ����ȷ����ϲ�������ɹ��ؽ������˺�С��i PSCs������Щ�о��л�õ�i PSCs��ʾ��������������û����ԴDNA�IJ��롣Ȼ����Ŀǰ�ķ����ϲ������崫�ݷ������ر��Ч�ʽ�����0.001%�����������ΪOSKM��˲ʱ���ﲻ���Բ�����ȫ�ı����Ŵ�����[14,36]��������ˣ��ٲ���������ת��ҽѧ�е�Ӧ�����кܴ��ǰ��[24]����һ�ַ�����ʹ�ø�����RNA��̨����(Sendaivirus,Se-V)����Ϊ�����������͵�ϸ������֯�е�����Դ����dz���Ч����Ҳ�����˵��ر��Ч�ʵ��ϰ�[39]��������ˣ���������Ŭ������һ�ָ�����Se-V[40]����ΪSe-V��������ά����������[41]�Ͱ��̲�����[42]������о��DZ����ϣ����������i PSCs��ϸ���������[43]��
����1.2.4 �������ϷDz�������ת��ϵͳ���ر��
����Ϊ�˲��������������ϵ�Ⱦɫ���i PSCs������ʹ��ϸ����RNA��������������(һ�����Ҹ��ƺ�ѡ�������)[44]���˳������DNA�Dz�������ϵͳ[45]������DZ�ܱ�ǻ���ֱ�Ӻ�˲ʱ�ش��ݵ���ϸ���С���Щ�����������ʹ�ã����ر��Ч�ʱ�LV��5��10��[24]����ˣ�ʹ��ϸ����RNA����DNA������Ҫ�㷺���Ż��Ա�������Ӧ��[24]��
����С����˳���άϸ���ѳɹ���ͨ��ֱ��ת�ƴ������ر�̵���[46]���ESCs[47]��ת����HEK293ϸ���з�����ܵ���[48]��ȡ������ر�̡�Ȼ�����ַ�������һ���ıˣ���Ϊ�����ϳ������ĵ����ʾ��кܴ����ս�ԣ���ת��Ч���ر�ͣ�ϸ���ر�̹�����Ҫ8�ܡ���ʵ����ָ��ͨ����ѧ�ر�̲���i PSCsҲ���ǿ��еģ�����һ���̿��ܵ���ͻ�䣬��Ϊϸ������������DNA���鵰������[14,33]������ϳ�RNA������ر�����ӵ���ʹRNA(m RNA)Ҳ���ǽ��������϶��ܸ�ϸ��������ƽ̨��������Щ����������Ҫ����תȾ���������ɾ��и���ȫ��i PSCs�������������[49]��
����1.3�� ����ر��Ч�ʵķ���
����Ϊ�˸Ľ��ر�̹��̣����Dz���������micro RNA(mi RNA)���·���������ر��Ч�ʡ����磬mi R-291-3p��mi R-294��mi R-295����������c-Myc�Բ������ȵ�hi PSCs����[50]�����⣬��ѧ������������ᡢ�����ƺ��鵰��������ø���Ƽ����ѱ�֤���ܴٽ�i PSCs������[51,52,53]�����������ĸı䣬��ȱ������Ҳ������ر��Ч��[54]������p53;��[55]������Nu RD(Mbd3/��С�����ܺ�ȥ��������������)���������ɳɷ�Mbd3�������Խ�һ���ٽ�i PSCs������[56]����ĸϸ�������������������ӣ���Glis1��H1foo��Ҳ������ر��Ч��[57,58]��
�������˿�������ر��Ч�ʣ���ЩС�������ر�̹����л��ܷ��Ӳ���OSKMת¼���ӵ����ã���������ȫ����OSKMת¼���ӽ���ϸ���ر��Ϊi PSCs�����磬ʹ���鵰��������ø���Ƽ���ת����������β���Ƽ���������������û��c-Myc��Klf4����������i PSCs�����ɣ����ҿ������Oct4��ά��ϸ���Ķ�����[59]�����⣬��Aza(DNA�������Ƽ�)��TSA(�鵰��������ø���Ƽ�)�յ����ܻ���Oct4��Nanog��Sox2�ı�����Խ�С�����ǰ��ϸ��(BPCs)�ر��Ϊi PSCs[60]����һ���о���������������ر��Ч�ʽϵͣ���ת��mi RNA-302/367���������û��ת��OSKMת¼���ӵ�����³ɹ��ؽ�С���������ϸ���ر��Ϊi PSCs[61]��Hou��[62]ʹ��7��С���ӻ�����ɹ���С����ϸ���ر�̳�i PSCs�����ر��Ч�ʸߴ�0.2%����Ȼ����С���Ӷ���ϸ�������ر�����ٴ����кܴ��Ӧ�ü�ֵ���������ر�̲���ȫ���յ���Ķ��ܸ�ϸ�����ָ���ϸ���������⽫����С�����ر�����ٴ��о��еķ�չ��
����2 ������i PSCs�ļ�����ģ
����2.1 ������༭����
��������i PSCs�ļ���ģ�͵Ľ�����ʹ��ͨ���������²��裺�ռ���Դ���ߵ���ϸ����ͨ���ر�̼��������ر�̳ɲ���������i PSCs������CRISPR/Cas9�Ȼ���༭���������Ȼ������(ͨ��ͨ���༭��֪������ػ���������Ľ���i PSCs)����ϸ���ֻ�Ϊ�����뼲����ص��ض�ϸ������ͨ���Ƚϻ����ͽ������ض�ϸ���������������弲�������Ա��ͣ��ڷ���ˮƽ���о���Щ���Ϳ���ʶ���µIJ������ƣ�Ϊҩ��ֺ��Ի������ṩ�µĻ��ᡣi PSCs�������ض�ϸ�����������ڼ�����ģ��ҩ��ֺ�ϸ�������е�Ӧ����ͼ2��ʾ��
�������ٷ�չ�Ļ�����༭���������ܹ����ض�λ�ý�����ı�����i PSCs����������������Դ��i PSCs�е��²�����ͻ���Լ����ض�ͻ������Ǽ���Ұ����i PSCs����Щ�����ṩ�������ͻ����ΪΨһ�ı�������������ƥ��ġ��Ȼ����i PSϸ��ϵ��ȷ���˶���ʵ�����Ŀɿ�������ͬʱ���������ڿ��ܵ�ϸ��ϵ����쵼�µ��Ŵ����������ż������Ļ������Ȼ���i PSCs������ģ��ɢ���Ի�����ʱ������Ҫ����Ϊ��Щ�����ı��Ͳ����С[63]��
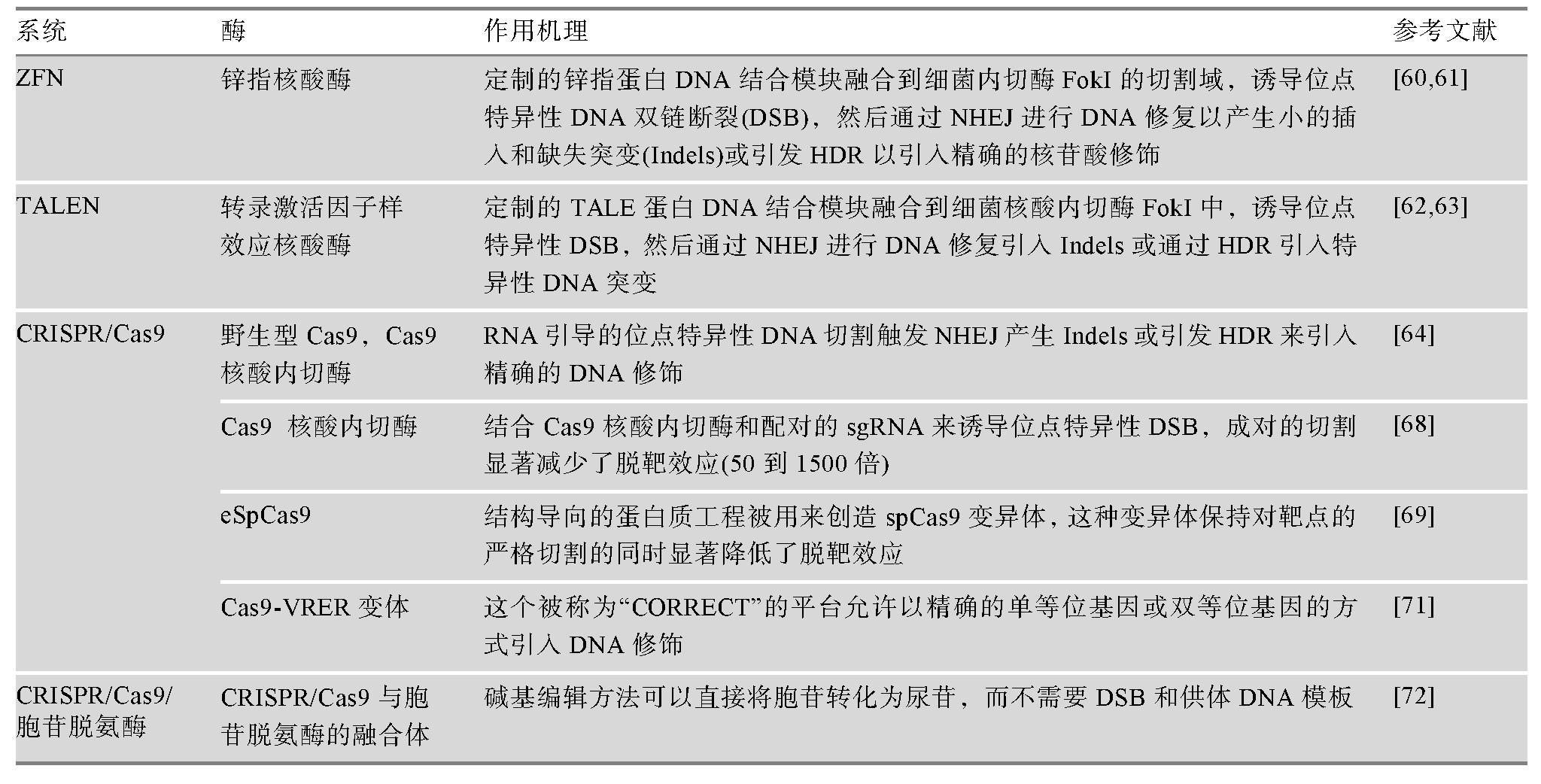
�����ɱ��λ�������Ժ���ø�Ŀ���������пָ����ø(zinc-finger nuclease,ZFN)[64,65]��ת¼��������ЧӦ������ø(transcription activator-like effector nucleases,TALENs)[66,67]��CRISPR/Cas9ϵͳ[68]��ͨ���յ���������λ���DNA˫�����ѣ������������ESCs��i PSCs�Ļ���༭Ч�ʡ�����CRISPR/Cas9����������Ƽ�ʹ�÷�����ŵ㣬��������̥��ϸ����i PSCs�Ļ���༭�еõ��˹㷺��Ӧ�á����ֻ���༭���������о���Ա���²�ͻ������ǻ���i PSCs����������i PSCs�е��²�ͻ�䣬�Ӷ���������i PSCs�ļ�����ģ�Ȼ������ƽ̨������ʹ��CRISPR/Cas9�������ܻ����ƫ��Ŀ���Ч������CRISPR/Cas9������ϸ��ϵ[69]�з�������Խϸ�ˮƽ�ķǰл������Σ������Զ��ʵ���ҵ�ȫ���������(whole genome sequencing,WGS)�о��������ǰл�����������������ϸ��(������i PSCs��ESCs)�к��ټ�[70,71]������༭����Ҳ�ڲ��ϸĽ������ƣ�����������ڽ���Ѱ�ЧӦ�����⡣���CRISPR/Cas9ͨ���յ�DNA˫���������༭һ��������λ�㣻�����ɳɶ���RNA����������ǿ�Ĺ��̻�Cas9����ø����(engineered Cas9 nuclease variants with enhanced specificity,e Sp Cas9)ָ����Cas9��������ø����(D10Aͻ����)Խ��Խ������ڻ�����༭[72]����Ϊ�����ַ��������ڱ��ֶе���ϸ��и��ͬʱ���Ž������Ѱ�ЧӦ[73]�����⣬��������Cas9 (dead Cas9,d Cas9)ͨ����ת¼����������������ںϣ����ڵ�����Դ�����ת¼��ͨ����ӫ�⵰���ں������������λ��[74]����CRISPR/Cas9ϵͳ�ĸ���Ҳ���Ծ�ȷ�ĵ���λ��˫��λ����ķ�ʽ��Ч������DNA���еı仯[75]������ڼ���༭�����һ���½�չ������CRISPR/Cas9������Ѱ�ø�ںϣ�ʹ����ֱ��ת��Ϊ���ն�����Ҫ˫��DNA����[76]�������·�������˻���༭Ч�ʣ�������һ���ٽ�����i PSCs�Ļ���༭��չ����������i PSCs�Ļ���༭�����Լ����û������1��ʾ��
����ͼ2 ��Դ��i PSCs���ض�ϸ�����������ڼ�����ģ��ҩ��ֺ�ϸ�������е�Ӧ��
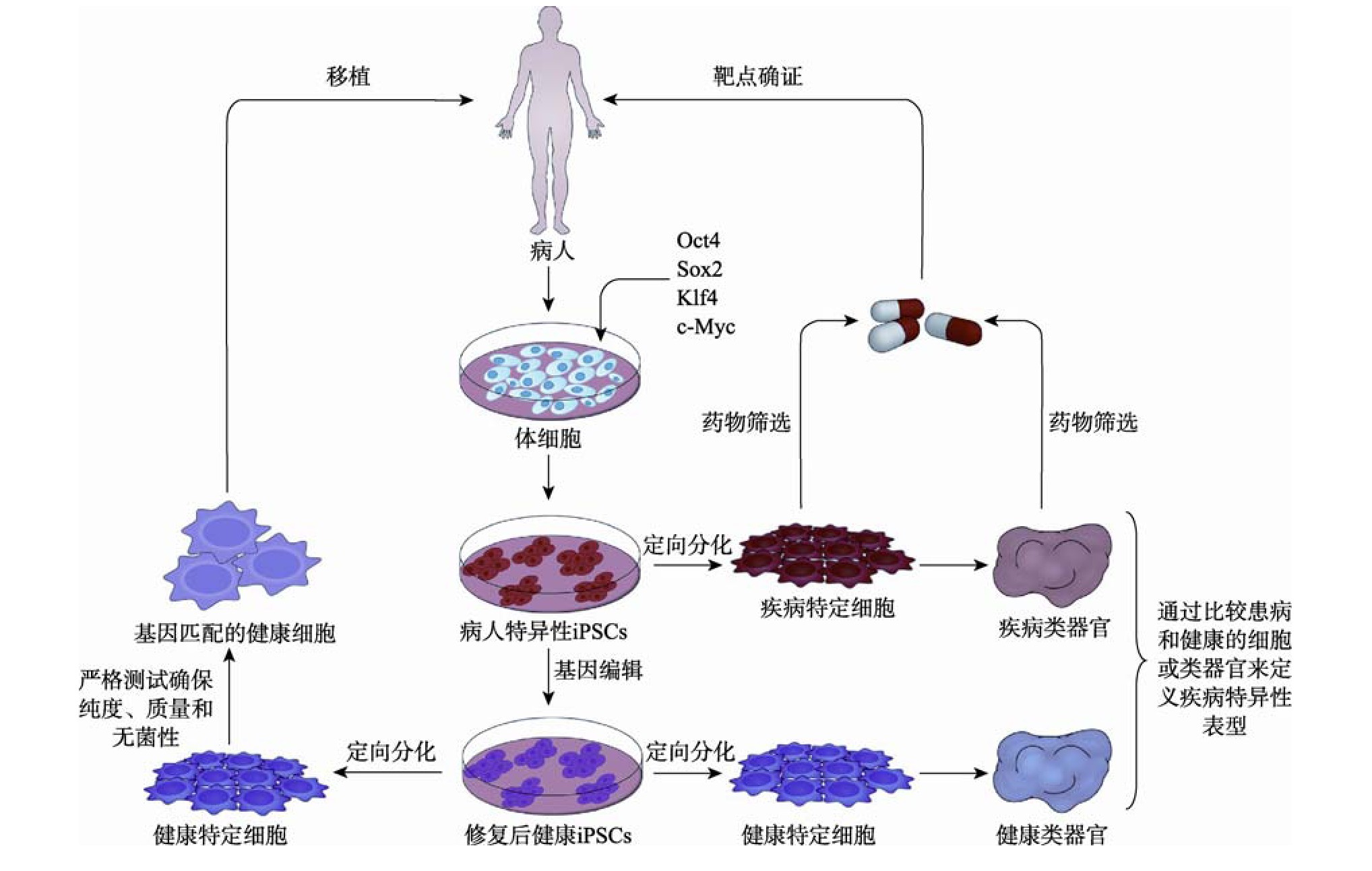
����Fig.2 Application of specific cells and organoids derived from i PSCs in disease modeling,drug discovery and cell therapy
����2.2 ����ͳ��άϸ����ģ
����ʶ�����༲���IJ������ƶ��ڷ����µ����Ʋ��Ծ��йؼ����á���Ȼ����ԭ��������Դϸ����������ģ���������о����༲���IJ�����ƶ����Ʋ��ԣ���һ���ؼ�����������������ÿ���չ�Ļ���ԭ��ϸ������������ϸ��������ϸ��������i PSCs�������Ҹ��µ��������Ժͼ������Էֻ�Ϊ�����κ�ϸ�����͵�DZ�ܶ���Ϊһ���������������Ʒ����Ϊ���༲��(�ر�����Щ����ȷ�Ŵ�ԭ��ļ���)ԭ���Ͽ���ʹ������õ�ϸ������(��Ƥ������άϸ����Ѫϸ��)��i PSCs��ģ������������i PSCs�����ṩ�����뼲����ص�ϸ��������ǰ����õ�ϸ�����ͣ�����Ԫ���ļ�ϸ�������⣬����i PSCs���Դ���ػ���������ã�������ǿ���ʹ���Ի�������ģ��Ϊ��ҽѧ�ĺ��IJ��֡�
��������i PSCs�ļ���ģ�ͱ��㷺�����о��ɵ�����ͻ������ļ���(������)[77]�����ַ����dz��ʺ��ڴ��༲������Ϊi PSCs���Ժ����ش���Щ�����������ϻ�ò��ֻ�������Ԫ���뼲����ص�ϸ�������磬�ӻ���i PSCs�зֻ���������Ԫ������ģ�⼹���Լ�ή��֢(spinal muscular atrophy,SMA)������һ����SMN1����ͻ��������緢�Լ�����SMN1����ͻ�䵼���˶���Ԫ���Ժ����ļ���ή��[77]������SMA����ͨ���ڳ�����6���³���֢״��������չѸ�٣�������ʱ����[78]��������Ļ���i PSCs�ļ���ģ���о���[77],i PSCs��Դ�ڢ���SMA���ߵij���άϸ�������ֻ�Ϊ�뼲����ص�ϸ������——�˶���Ԫ����δ��Ӱ��Ķ�������ȣ�����i PSCs�ֻ����˶���Ԫ����ʽ��͡����⣬��Դ��SMA���ߵ�i PSCs�ܹ��Ա�������ײ�ù��(��֪���ֻ�����������SMN���ı�����)������Ӧ���������ֻ�����������SMA����i PSCs��SMN����ˮƽ[77]�������о��ṩ��һ��ԭ���Ե�֤�ݣ�������Դ��i PSCs��������ģ���緢���Ŵ�������������ΪDZ�ڵ�ҩ��ɸѡƽ̨��
������1 ��������i PSCs�Ļ���༭����
�����Է��������ļ������н�ģ������ս�ԣ���Ϊ������i PSCs�ֻ�������ϸ��ͨ�����ֳ�������ı��͡����磬�����������Դ��i PSCs���ļ�ϸ������������ҽѧ��ҩ���ʱӦ���dz���ģ������Ƚ������ڳ����ļ����ļ�ϸ���ij���ȣ��Ӷ�ʹ����ϸ����ʾ�����Ƶ������ԡ����������ܺͶ�ҩ��̼��ķ�Ӧ��Ȼ��������ʵ�У�i PSCs��Դ���ļ�ϸ����δ����ģ�����̬�����ȷ����������̥״̬�µ��ļ�ϸ��[79]��δ�����ļ�ϸ��������ʾ����֯�ļ��ڽṹ�ƴ������ƣ���Щ������ӳ�ڳ�����ؼ��ڻ���(��MYL2��MYH7��TCAP��MYOM2)������ת����ػ���(��KCNJ2��RYR2)�ĵͱ�����[80]���о���Ա�����յ��ļ�ϸ�����췽�����˺ܶ�Ŭ�������Ӽ�״�ټ���[81]����������[82]������ά�ļ���֯�еĻ�е���ڽ�ϵ�̼�[83]�ȷ�������i PSCs�����ļ�ϸ���ij��������á�����Ҫ���i PSCs����ϸ����������͵������⣬��ģ�����Լ��������������������յ�ϸ��˥�ϡ�����i PSCs������ϸ�������յ�˥�ϵ�һ�ַ�������ϸ��Ӧ��Դ����ϸ�����������������غ�MG-132�ȵİ��������幦�ܻ��ʽ���Ļ�����[84]����һ���յ�ϸ��˥�ϵķ�������λ��������˥�����յ���˥�Ļ������[85]��Ȼ����ϸ��Ӧ��Դ����˥���ı����Ƿ���ͨ��һ������������˥�ϵĻ����յ�ϸ��˥�����д�ȷ����
����i PSCs��Ϊ�о�ɢ���Լ����ṩ��һ���µķ���(�䲡����δ�ڻ��ߵļ���ʷ�����ͻ����ȷ��)����һ��ʮ����Ҫ����Ϊ���༲���Ĵ�������߶�����ɢ���Լ��������磬������մ�֢�У�95%�IJ�������ɢ���Լ������о���Աͨ����ɢ�������ĺ�Ĭ������i PSCs������ϸ���ķ������֣�һЩɢ���Բ������ֳ����ض�����ͻ��ļ��������ĺ�Ĭ����ͬ�ı���[86]�������ʹ��i PSCsҲ�����Զ�ɢ���Լ����������¼�����Ȼ����ʹ��i PSCs��ɢ���Լ������н�ģͨ���ȵ����������ѣ���Ϊ���༲���ı��ͱ仯���ɶ��СЧӦ�Ŵ����ձ�����ͻ������ع�ͬ�յ��ġ���Ȼ��Դ�ڴ��༲�����ߵ�i PSCs���ܰ����뼲����صķ��ձ����壬��ʹ��i PSCs��ģ����༲�������Ŵ��ͱ����Ŵ����������б������ø��ӣ�����ɢ���Լ���i PSCs������ϸ���ı���Ԥ�Ʊȵ�����i PSCs������ϸ����Ϊ��ϸ����ˣ�����i PSCs��ɢ���Լ���ģ�͵�һ���ؼ���������β���������ط��ձ����ϴ��ڲ���ijɶԵȻ���ϸ��ϵ[63]������CRISPR/Cas9������������༭�ĵȻ���i PSϸ��ϵ�Ĺ��ܿ��Դ���һ�����õĿ���ϵͳ��ȷ���뼲����صĻ�����ձ������ΪΨһ�ı���[87]�����ַ��������ڲ�������ɭ����ͬ��ط��ձ�����ĵȻ���i PSϸ��ϵ����ϵ�λ���������Է����ܹ��Ը��Ŵ����ձ��������ǿ�����ķֱ�ͻ���λ[87]����һʵ����Կ������о�������������ص��Ŵ�Σ�����ء�
�������⣬����i PSCs�ļ���ģ��Ҳ��Ϊ�о����������ķ������ƺ�ɸѡ�µ�����ҩ������ù��ߡ��ڲ��ı�ϸ�����������е�����£���Դ�������������ߵ��κ�ϸ�������ر�̳�i PSCs���ٷֻ�Ϊ���Ӧ����������ص�ϸ�����ͣ��Դ��о�i PSCs������������ϸ��ת���Ĺ��̡����磬��������赥��ϸ����Ѫ��(juvenile myelomonocytic leukemia,JMML)��һ����Ϯ�Թ����������������䷢����ԭ����ͻ�䵼��ϸ�����������ź�ת���쳣��������JMML���ߵĹ���ϸ��������Ѫϸ���ر�̳�i PSCs����i PSCs�����뻼����ϸ����ͬ��PTPN11�����p.E76K����ͻ�䣬��ͻ�������SHP-2 (һ�ַ������Ұ�������ø)[88]��������i PSCs����ֻ���������ϵϸ����ȣ�JMML-i PSCs��������ϵϸ����ֳ������ǿ����ϸ��–����ϸ������̼�����(GM-CSF)��������STAT5/ERK���ữ��ǿ����Щ������JMML���ߵ�ԭ����ϵϸ�����ƣ���MEK��ø��ҩ�����������ܽ���JMML-i PSCs��������ϵϸ��GM-CSF�Ļ���[88]�������о�����i PSCs����ϸ��������ԭ���Զ����������⽨ģ�е�Ч�ã�Ϊ�ü���DZ�ڵİ��������ṩ�������ݡ�����i PSCs�ļ�����ģ���Ŵ��������������о���Ҳչʾ�����ǰ����Li-Fraumeni�ۺ���(Li-Fraumeni syndrome,LFS)��һ�ֳ�������p53����ͻ������Ķ��������ۺ���[89]����Դ��LFS���ߵ�i PSCs�����ijɹ�ϸ����ʾ���ֻ�ȱ���Ժ�������,��H19����ı������裬��Щ�����������(osteosarcoma,OS)ϸ��һ�¡��ָ�LFS-i PSCs��H19�ı���ٽ��˳ɹ�ϸ���ķֻ�����������������[90]�����о����֤������i PSCs�о��Ŵ����������������Ŀ����ԡ�
����2.3 �������٣���������ĸ�����֯
����i PSCs������Ʒ���ٴ�Ӧ���ںܴ�̶��������ڶ���ֻ���ϸ��״̬ת������֯���̵����¼�������������i PSCs����ֻ����о��������õ������е�ϸ��ˮƽ���ͣ���ת��Ϊ��֯ˮƽ������ˮƽ�ļ�����Ҫ���������ӵ�3D��ϸ��ϵͳ������i PSCs���������ٵķ�չ��i PSCs�ڼ�����ģ����ȡ������Ҫ��չ[91]�����������ɸ�ϸ���ֻ���������ά��ϸ���ۼ��壬�ܹ�������֯�����Ա��ֳ�����֯�Ľṹ������ϸ����������[92]������ָ����������i PSCs�ֻ��Ŀ�������������ѧ�����������ƣ��Բ���Խ��Խ���ӵ�“������֯”[91]��������Դ��i PSCs�ͻ��������i PSCsƽ�зֻ��������٣����Խ�������ˮƽ�ļ������������ض��ķ��Ӳ��䡣һ��������������������ˮƽ�о�ƽ̨������������پͿ�������ҩ��ɸѡ����֤�о���
�������ڵ�ESCs��i PSCs�о������ֻ�������ģ����ϵͳ����[93]������i PSCs������3D���������ٵĴ������ǽ�������Щ�������ֻ������Ļ����ϣ������Ҫ���������ﷴӦ������������70�죬������Щ3D���������ٻ����о���Ա������̬���ɵ���������[94]����Щ���ٰ�����Ƥ����֯�����Լ��ض�ǰ�Ժͺ�������Ĺ�������Ԫ����������δ��������Ĥ������Եķֻ��ṹ[94]��Quadrato��Sloan��[95,96]��3D�������������������ĸĽ������յ�3D�������γɴ����еĺ������С�Ե��ض�����ṹ���Լ��յ��γ�Ƥ���۵��ṹ������������Ϊ���ŵĸ����Զ�������ģ����ֵ�����Ͷ������ϵͳ��������Щ�������о�Ϊ�����IJ�������ѧ�ṩ�����Ļ��������磬CDK5RAP2����һ�������嵰�ף��õ�������˿���ѹ����ж�λ�ڷĴ��弫����CDK5RAP2����ĸ����Ӻ���ͻ�佫����Сͷ���Ρ�CDK5RAP2����ͻ�仼��i PSCs�����������������������ģ��Сͷ���Σ��ɴ˷��ֻ������е��������پ���������Ƥ���ڷֻ�������ʾ���쳣�ľ�����ȡ��ͽ�С�ķֻ�����֯����[94]������һ�ֱ��������嵰��CENPJ����������װ�ͳɺ˵ĵ���������Ҫ���ã�CENPJ��ͻ�佫����Сͷ�ۺ������ò�����i PSCs����������������Ҳ�۲쵽���Ƶı���[97]������������Դ����i PSCs���������ٻ��������о���������ƽ̨���о���Աʹ�ú�����ϸ����ǰ��ϸ���Ĵ��������٣�ͨ������ת�������볣������������ͻ�䣬Ȼ���о�ͻ���ϸ���Ĺ�������������ģ�������ķ����ͷ�չ[98]����ˣ���2Dϵͳ��ȣ��������ٵ��о���������ǿ�˶���ϵͳ������������ѧ���˽⡣
�����ڴ�i PSCs���ɸ�ϸ�����Ƹ��ָ��༲������Ķ�ά�ֻ������ѱ�֤������Ч��[99]�����������ά������ϵͳ�У��ӻ��иε�������i PSCs���Ѿ�����������ϸ����Ŀǰ�ķֻ����������ȶ���ֻ�Ϊ��Ƥϸ����ĸϸ����Ȼ������ά�����зֻ�Ϊ����ϸ����ϸ��[100,101]����Щ����ϸ�������ٿ���������֭�Ტ���й����Է������ã��Ӷ�����ģ��Alagille�ۺ���������һ������Notch�ź��ж϶����µ����γ�����ļ���[101]����������Ըβ����߷ֻ��ĵ���ϸ���������У��ϳ�������������������Ľ����������ٵĴ�С�����ҩ�������ƻ��߶����Ը��״��е�����һ��[101]��Я��F508delͻ��Ķ����Ըβ���������ά��(CF)���ߵ�i PSCs�����ĵ���ϸ����������ʾ���Ȼ���ת�������CFTR���ױ��������ͣ���CFУ��ҩ��VX809���ȶ�����������CFTR���ı�����ҩ����������ά�������е�����һ��[100,101]����ÿһ�������У����Ӽ������͵Ľ�ģ����ͨ����ά�����ٵķ�չ��ʵ�ֵġ�
���������������������ٵ��о�ȡ���˼���Ľ�չ��i PSCs���������������پ���̥�����ֻ����ص㣬���ѱ������ļ�ϸ�����˺�������ģ���о���������һ����ǿ����һ�ֹ۵㣬����Դ��i PSCs���ļ�ϸ����������֯�������ֳ�̥�����ֻ�������ܻ�������˼����Ľ�ģ[102,103]��������о��߲����˾������Ŀ�ǻ�ĸ����ļ�ϸ�������٣����÷ֻ�����������[104]�����⣬i PSCs�Ѿ������ڹ�����ά�ļ���֯����������е������л��ϸ������ʶ��ļ�ϸ�������Ӱ�죬��Щ�о�����ʹ��i PSCs����֯���̵ĸ�������[105]���о���Ա��ͨ��3D����оƬ������i PSCs����ģ���ļ�������һ���Ա�Ӧ����������������ϵͳ�У�Ϊģ��Ѫ�ܹ�ע�ṩ�˶��������֧��[106,107]��
��������������о�ǿ������i PSCs�����ķ��������ڼ�����ģ�е�����[108,109]�������ǽ���i PSCs����ֻ�ΪNKX2-1+��������ϸ��������ϸ���ܹ�����γɽ��˻�Զ������ϸ��[108]���ڵ�Wnt�������ά�����У�NKX2-1+��ϸ�����ظ��γɰ������ֽ�������ϸ���������٣���������ϸ������״ϸ���ͻ���ϸ��[108]����forskolinͬʱ��������������ά��F508del-CFTRͻ�䴿���ӻ��ߺ����Զ��յĽ��������i PSCs�����Ľ������������٣����֮�£�������������������ʾforskolin�յ�����������[108]���о���Ա����һ��������CFTR�����ϰ�������i PSCs��F508del�Ļ������������forskolin�յ�������[108]��������ʾ�˻���༭��i PSCs����ģ����ȷ�ϵĻ�����-����ϵ����i PSCs��Դ��NKX2-1+����ϸ����3D������Ҳ�ֻܷ�ΪԶ����������٣���Щ���������ٺ��й����Ե�2�ͷ�����Ƥϸ������Щϸ�����а��С�岢���ڱ�����Ե��ס�������ȱ��������Ե���B (SFTPB)���ߵ�i PSCs���������ķ�����������Ȼ����2�ͷ�����Ƥϸ��������Щϸ��ȱ�ٰ��С����ܺϳ�SFTPB[109]�������о��������ͨ������༭������SFTPBͻ��Ҳ���ܸı�ü�������i PSCs����ϸ���ı��͡�
����2.4�� ��–����Ƕ����
�������ܻ��������ٵļ���ģ���ڸ����ԡ�����Ⱥ�ϸ�������Է���������չ�������Ǿ�������֯������������������ѭ����������ϵͳ�������Լ�������л��������õ�̽��[110]�����⣬�����ٻ�ȱ�����ڴ��ڵ�������̬�����ź�[92]�����磬��ģ��ѪҺ����ʱ������ֻ�ϵͳ���ٽ���������������Ѫ�ֻ�����Ѫ��ϸ��(HSCs)���о���[111,112]����������ϸ����������������ֲ����i PSCs���о���չ�Ѿ��˷�����Щ�����ԡ������������ϸ�����Ƶ��ٴ��о������ʱ�ը������֮�ƣ�������i PSCs��������ֲǶ���弲��ģ���Ѿ��ܹ�������ʵ��ģ�����༲������Ϊ�ƶ���Щ�о���չ�Ŀɿ����ߡ�
������������֯��������ֲ�Ѿ������о����༲����ʮ���ˣ�����̥��ϸ���ķ���������i PSCs�����ķ�չ������̥��ϸ�����ֻ������Լ���ֲģ�������Ϊϸ�������ṩ��һ�����͵����Ƽ���[113]��������i PSCs�ֻ�����Ԫ������ֲ�������е�С������ʱ�������ϡ��γɹ�����ͻ�������ϵ���·��[114]��i PSCs�ֻ�����Դ����ϸ��(NSCs)�ڴ�������ֲ���ɷֻ�Ϊ��Ԫ�����ν���ϸ�������ɴٽ�ȱѪ������ģ�͵Ļָ�[115]���������������鳤�ද��i PSCs�зֻ������Ķ�Ͱ�����Ԫ����ֲ�����Ĵ��ԣ���������ɭ��ģ�͵Ĺ��ܣ��������鳤��i PSCs�Ķ�Ͱ�����ԪҲ����������ֲ���������ڳ���ֲ��[116]�����������ŷ��Ĺ��ָܻ����ٴ�ǰ֤��Ϊ��������ɭ�����ٴ������ṩ��ǿ������֤����������ϸ���Ʒ����������ڵ������ӣ�Ŀǰ������Ϊ�о��������֢�����Ʒ���[117]�����������Ѫ��ϵͳ��ע����ĵ�����ѧʵ�������˽���i PSCs�������������ٳɹ�ֲ�뵽����С�����У���Ϊ��ϵͳ������ȫ��λ��ģ�ṩ��һ����ƽ̨[118]����֮����Щʵ��֤����Ƕ��ģ������i PSCsģ��������ϵͳ�����еļ�ֵ�Լ�����i PSCs��ϸ����������Щ�����еĿ����ԡ�
�����о���Ա�Ѿ�����i PSCs��������������ֲ��С�����ڣ�������Ѫ��ϵͳ���о����������������������[119]����Դ����i PSCs�ĸ����߲�ϸ����������Ƥ��ϸ���ͼ���ʸ�ϸ�������������γ�ԭʼ�ĸ�ѿ����Щ��ѿ������ֲ����С��Ѫ��ϵͳ�γɹ�����ϵ��Ȼ���˰����ͷŵ�С��Ѫ���в������������е�ҩ���л����[119]���о������������ֲ��Щ��ѿ�������С���˥��ģ�͵Ĵ����[119]���Ӽ����Ըߵ��̴�Ѫ֢���ߵ�i PSCs�зֻ����ĸ�ϸ���Ѿ���������ֲ��ģ�����ּ���[120]��
����������������ֲ������ȱ��С��ģ�����ǽ����������о���һ��ؼ�������������i PSCs����ѪҺ����ģ���У��Ѿ��������γ���ѪǶ����İ���[121]��������ϵ��Ѫ��(acute myeloid leukaemia,AML)ϸ����MLL(Ҳ��ΪKMT2A)�������Ϸ�����λ������ͨ������쳣��MLL������DNA��������Ч�ؽ������±��Ϊ������ϸ���������ٷֻ�Ϊ����Ѫϵϸ��[121]��Ȼ�����ڶ���ֻ�Ϊ��Ѫ��ϸ������Щϸ������������ֲ�����б��ֳ��쳣�����Ҹ�����������˸�ϵͳ������ģ����AML�Ŀ�¡�ṹ����������������Щi PSCs�����ǿ�¡��ҩ�����еIJ�����[121]����������i PSCs��Ƕ����Ҳ������ģ��ʵ���������˽���ĸϸ����ϸ���ر�̳�i PSCs�������������������ӵĸ������ܣ���i PSCs����ֻ�Ϊ����ϸ��ʱ����Щϸ�������˶���DZ�ܣ���������ֲ��С����״��ʱ�γ���Ϯ������[122]���ӻ���Li–Fraumeni�ۺ����Ļ����л�õ�i PSCs�����ijɹ�ϸ��Я������ϵTP53ͻ�䣬��Щ����������ʵ�����������ѪҺϵͳ����������������ȱ��С��Ƥ��ע����γɹ�����������[123]�����������Դ��i PSCs�������ٺ�������ֲ��С�����ڵ��о�����Ϊʵ�����ķ����ͷ�չ�ṩ�µ��ӽǡ�
����4 ������i PSCs���ٴ��о�
�������˼�����ģ֮�⣬i PSCs����DZ��֮һ�Ƕ������Ŵ��������Լ�������ϸ���ͻ����������[124]�������ַ����У����˵���ϸ�����������������ͨ��������Dz����鵼�Ļ���ת�Ʊ����±�̳�i PSCs������ʹ�û���༭������ת�������Ի�����Դ��i PSCs���л���������������ɸѡ����i PSCs�ֻ�Ϊ��Ӱ���ϸ�����͡��ڽ���ϸ�������Լ����ȡ����ԺͰ�ȫ�Ե������������������ƥ��Ľ���ϸ����ֲ���������ڽ���ϸ�����ơ�����������ֲ�ķ������Է�ֹ���صIJ���֢����ͨ��������ͬ��������ֲ�����ֲ���������i PSCs�ڼ��������е�Ӧ����ͼ3��ʾ��
����Ȼ�����ڳ�����ٴ�Ӧ�ÿ�ʼ֮ǰ������iPSCs�������м����ϰ���Ҫ���������һ����iPSCs����������[125]������iPSCs���������������˺ܳ�ʱ�䣬���ǻ���ۺ����쳣��������������Ӻ���ɥʧ[126]����ˣ����ٴ�ʹ��֮ǰ��iPSCs������Ʒ��Ҫ��ϸɸѡ��ȷ���Ƿ����DZ�ڷ��յĻ���ı�[126]���������ϸ������ȷ���䴿�ȡ����������ԡ���һ�����������ɿ������ٴ���iPSCs�ijɱ��ܸ��Һ�ʱ�ϳ���������������Ҫ�����ܹ�������������ϸ����ԭ(HLA)���ϳ�����iPSCs��ϸ����[127]������һ��������2013���ձ������˾�����ѧiPSCs�о���Ӧ������(CiRA)��CiRA��һ�������iPSCs�⣬�ռ���50�����Խ���־Ը�ߵ�����ѪT�ܰ�ϸ������ѪHLA���ϱ걾����HLA��������Ҫλ��(HLA-A��HLA-B��DRB1)����ƥ��[128]��2015�꣬Cellular Dynamic International,Inc.��������һ��ϸ�����ƿ⣬��������19%�������˿������������ţ��Կɳ��ܵijɱ�Ϊ���������ṩ��������ϸ����HLA�����ʹ���iPSϸ���������һ����Ч��ϸ�����ƵIJ��ԡ��������ϰ���iPSCs����ϸ���������ԺͲ������ԡ�ͨ����iPSCs������ϸ�������ʵġ����磬��iPSCs������Ԫϸ���ĸ��ַ����У�����ϸ������������ͬ���͵���Ԫ������ϸ��(�罺��ϸ��)[129]�����ٴ�ʹ��ǰ�������ϸ�������������ԣ���ȷ��ֲ��ϸ����һ���ԡ���ˣ�����ͨ��ӫ��ϸ����ѡ��(fluorescence-activated cell sorting,FACS)��ż���ϸ����ѡ��(magneticactivated cell sorting,MACS)��iPSCs����ϸ�����з���[130]����ѡ���ٴ��о���Ҫ���������ϸ����������iPSCs�ֻ����ĸ���ϸ������ʾ��������̥������ϸ���ı��ͣ������iPSCs������ϸ���Dz�����ġ����Ӽ�״�ټ���[81]����������[82]�ȷ������ܴٽ�iPSCs����ϸ�����죬��iPSCs��������֯��ΪǶ����ֲ�붯�����ڽ���һ���ٽ�iPSCs��������֯����[131]����iPSCs����ϸ����Ȼ�����ϸ����״̬��һ����ࡣ������ֲ�������ټ����Ľ�һ����չ�������ڿ˷�����ϰ��������ٽ�iPSCs�ٴ��о��ķ�չ��
����ͼ3 iPSCs�ڼ��������е�Ӧ��
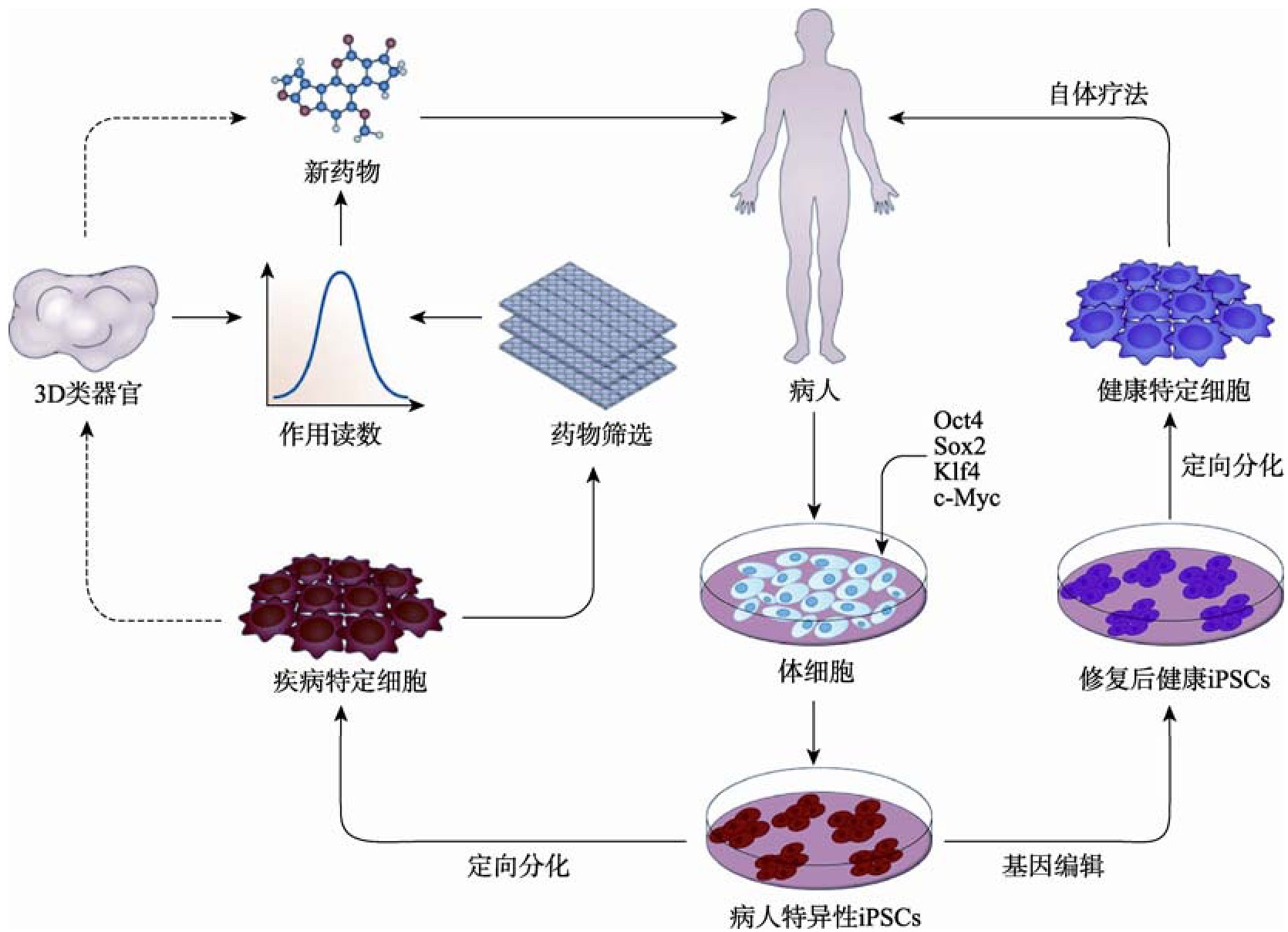
����Fig.3 Application of iPSCs in disease treatment
������һ��i PSCs�ٴ�����(��һ��)��2014��9�����ձ����С�����RIKEN��չ����ѧ���ĵĸ����Ŵ�֤���˻���i PSCs������Ĥɫ�ر���(RP)���Ƶİ�ȫ�ԣ�RP����Ҫ�������۾��ڶ����Ѫ���γɵ���������ȫɥʧ�������ڽ�������i PSCs����������Ĥɫ����Ƥϸ����ֲ���ƺ�,������������[132]������෴���ǣ�Kuriyan��[133]������ʹ������֬����ϸ�����������Իư߱���(AMD)�������Խ��������������Ѹ��ɥʧ����Ҫ������������Ϊ����Ҫ�������Ʊ�ϸ����ʹ������i PSCs��Ʒ���и��Ի�ҩ�������ƺ��ǵ�����������ѡ�����ڸ������ļ������ر������ļ������ȼ��Գ�������������i PSCs�Ʒ����ܲ������ڴֻ��ߣ���Ϊ��ϸ����ÿ��ϸ��ϵ��Ҫ�߰��ijɱ��ʹ�����ʱ�䡣Ҳ��Ϊ��Щԭ���ձ�i PSC-RPEʵ��ĵڶ��ν�ʹ��������Ʒ[134]����Щ�¼������˻���i PSCs��ϸ�����ƻ��кܳ���һ��·Ҫ�ߡ�
����Ƕ�Ͽ�ԭ����(chimeric antigen receptor,CAR)Tϸ�����Ʋ����ѱ�֤���ܳɹ��ؽ鵼ѪҺ�������������ˡ��������Ʊ�����Tϸ�����ܳ�Ϊ�������ر��������������ƻ�ϸ���������ơ�������ܰ�ϸ�������ͽ���T�ܰ�ϸ������Ч����[135]����������ǶԻ���������i PSCs���и��죬�Ӷ����ɺ�HLA�صĿɶ��ƿ�ԭʶ��Ŀ�ԭ�����Ե�CARTϸ������������༭����ȫ��ͬ������i PSCs (������HLA���������)���Խ�һ���ֻ�Ϊȫ���ܵ���֯��������������Tϸ��[136]������Ϊֹ������ģ�͵��ٴ�ǰ�о�������i PSCs�����Ĺ���CARTϸ��(i CART)ϸ�����п���������[137]��2019�꣬Ci RA��Takeda������������һ��i CARTϸ�����Ƽƻ�����2021�꿪ʼ�ٴ������һ�Ρ�i CART���ƿ��Ը�����ҪΪ�������������ֳɵ������Ʒ�[138]��i CARTɱ����ϸ����DZ���洦�������ٴ�����������Ӧ�õ�һ����Ҫ��̱���
������Tϸ����ȣ���Ȼɱ��ϸ��(NKϸ��)��һ���ܰ�ϸ��������Ȼ����ϵͳ�鵼�������Ϳ��������Ե������з����Źؼ����ö�����MHC���ơ�Ӧ��NKϸ�������Ʒ����Ƽ�����ϵ��Ѫ�����ٴ���������ʾ�����ŵ���Ч����������������������Ч��ϵ�[139]����NKϸ����Tϸ���Ľ���ϸ��Ⱥ�����ѻ�ã�ͨ���ɵ���ϸ��������Ѫϸ�������ʻ������ɡ�i PSCs�ij����ṩ��һ�ֶ��صĽ����������������ͬ�ʺ���ȷ��NKϸ��Ⱥ����ЩNKϸ��������ͨ�������������߿��������ԣ������ٴ����ģ����“�ֳ�”ϸ��[140]�����κ��NKϸ�����Ա���������壬�������DC16-Fc�����CAR���壬Ҳ�����������Ʒ�����Ӧ��������俹ʵ����������[141]����ĿǰΪֹ����һ���о���ʾi PSCs��Ϊһ��ƽ̨�������а���������CAR�����NKϸ���Ŀ����Ժ���Ч�ԣ����ѳ���������ֲģ������ʾ������ӡ����̵Ŀ��������ԣ��Ҹ�ϸ�������ڵĴ����չ���Խ�С[142]��2018��11�£������ٴ�����(NCT0384110)�������ʳƷ��ҩ�������(FDA)�����״�ʹ�ÿ�¡i PSCs�������ֳ�NKϸ��(����ΪFT500)��Tϸ��Эͬ���ã��Ը��õ�����ʵ��������������������߸�ҩ(ÿ�θ�ҩ��1×108ϸ��������ϼ������Ƽ��͵���¡����)��������ȫ��������ʾ�����28��Ĺ۲�����û�����ز����¼���2019��2�£�FT500���Ʒ�����64����ͬ�������������н����˽�һ���İ�ȫ�Բ��ԣ��ٴ�����Ŀǰ���ڽ�����(NCT0384110)���ձ�������ѧ����Ȩ������ҩ(Takeda Pharmaceuticals)��������i PSCs���ֳ�CARTϸ��ƽ̨��������ҵ������һ���ٴ������Ŀ������Ϊ2021�ꡣ����Щ�ٴ�ǰ�о�ת��Ϊ�ٴ������DZ���������ڴ��ġ�
����5 ��������չ��
������i PSCs�������������������������о��������ƺ�DZ�����Ʒ���ͨ��������i PSCs�������¼������϶�ȡ���˺ܴ��չ�������м�����Ҫ�����д��������i PSCs���������ļ�ϸ������ϸ���ȸ���ϸ������ʾ������ȵIJ����⣬i PSCs���ر�̹�������ϸ�����ԡ����䡢��л��ص���ϸ������Ĵ���Ҳ��������עĿ�����ǵ�i PSCs���ܱ���������ϸ��������ص���ϸ������(��������Warburg��л��)[143,144]��ɸѡ��������Щ�����Լ�����ֲʱ�����γɵķ��գ������о���Ϊ�����������ش���Ŀǰ��ȱ����Ч�ķ����������������Щ��ϸ�����䡣
��������������i PSCs�����ķ�չΪ��������Ƽ����ṩ��һ��ǿ�������·�������Ϊi PSCs������һ�ַ�ʽ��ת�䣺��������ֱ�ӹ۲��������صIJ���ϸ������ʾ�˻��������¹�ϵ���ؿ��ͼ��������ǶԸ��ּ�����չ�����⡣��CRISPR/Cas9��3D�����١�����Ƕ�����micro RNA���ӿ��ص�Ϊ�������¼����ij��ֺͲ��ϸ��£�����һ���ƶ�����i PSCs�ļ�����ģ�����Ʒ�չ����Ȼi PSCs��Ӧ���д����Σ�Ӧ�÷��սϴ���������һ��15�꣬i PSCs��Ӧ�û��ڸ��廯����ҽѧ�������������Ʒ���չ�ֳ�������ǰ����
���������
����[1] Waddington CH.The strategy of the genes.Allen and Unwin:London,1957,Reprinted 2014.
����[2] Gurdon JB,Elsdale TR,Fischberg M.Sexually mature inpiduals of Xenopus laevis from the transplantation of single somatic nuclei.Nature,1958,182(4627):64-65.
����[3] Wilmut I,Schnieke AE,Mc Whir J,Kind AJ,Campbell KH.Viable offspring derived from fetal and adult mammalian cells.Nature,1997,385(6619):810-813.
����[4] Evans MJ,Kaufman MH.Establishment in culture of pluripotential cells from mouse embryos.Nature,1981,292(5819):154-156.
����[5] Thomson JA,Itskovitz-Eldor J,Shapiro SS,Waknitz MA,Swiergiel JJ,Marshall VS,Jones JM.Embryonic stem cell lines derived from human blastocysts.Science,1998,282(5391):1145-1147.
����[6] Davis RL,Weintraub H,Lassar AB.Expression of a single transfected c DNA converts fibroblasts to myoblasts.Cell,1987,51(6):987-1000.
����[7] Takahashi K,Yamanaka S.Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors.Cell,2006,126(4):663-676.
����[8] Takahashi K,Tanabe K,Ohnuki M,Narita M,Ichisaka T,Tomoda K,Yamanaka S.Induction of pluripotent stem cells from adult human fibroblasts by defined factors.Cell,2007,131(5):861-872.
����[9] Yu JY,Vodyanik MA,Smuga-Otto K,AntosiewiczBourget J,Frane JL,Tian SL,Nie J,Jonsdottir GA,Ruotti V,Stewart R,Slukvin II,Thomson JA.Induced pluripotent stem cell lines derived from human somatic cells.Science,2007,318(5858):1917-1920.
����[10] Masui S,Nakatake Y,Toyooka Y,Shimosato D,Yagi R,Takahashi K,Okochi H,Okuda A,Matoba R,Sharov AA,Ko MS,Niwa H.Pluripotency governed by Sox2via regulation of Oct3/4 expression in mouse embryonic stem cells.Nat Cell Biol,2007,9(6):625-635.
����[11] Nakagawa M,Takizawa N,Narita M,Ichisaka T,Yamanaka S.Promotion of direct reprogramming by transformation-deficient Myc.Proc Natl Acad Sci USA,2010,107(32):14152-14157.
����[12] Niwa H,Ogawa K,Shimosato D,Adachi K.A parallel circuit of LIF signalling pathways maintains pluripotency of mouse ES cells.Nature,2009,460(7251):118-122.
����[13] Takahashi K,Yamanaka S.A decade of transcription factor-mediated reprogramming to pluripotency.Nat Rev Mol Cell Biol,2016,17(3):183-193.
����[14] Stadtfeld M,Hochedlinger K.Induced pluripotency:history,mechanisms,and application.Genes Dev,2010,24(20):2239-2263.
����[15] Soufi A,Donahue G,Zaret KS.Facilitators and impediments of the pluripotency reprogramming factors’initial engagement with the genome.Cell,2012,151(5):994-1004.
����[16] Papp B,Plath K.Epigenetics of reprogramming to induced pluripotency.Cell,2013,152(6):1324-1343.
����[17] Plath K,Lowry WE.Progress in understanding reprogramming to the induced pluripotent state.Nat Rev Genet,2011,12(4):253-265.
����[18] Nakatake Y,Fukui N,Iwamatsu Y,Masui S,Takahashi K,Yagi R,Yagi K,Miyazaki JI,Matoba R,Ko MSH,Niwa H.Klf4 cooperates with Oct3/4 and Sox2 to activate the Lefty1 core promoter in embryonic stem cells.Mol Cell Biol,2006,26(20):7772-7782.
����[19] J?hner D,Stuhlmann H,Stewart CL,Harbers K,L?hler J,Simon I,Jaenisch R.De novo methylation and expression of retroviral genomes during mouse embryogenesis.Nature,1982,298(5875):623-628.
����[20] Matsui T,Leung D,Miyashita H,Maksakova IA,Miyachi H,Kimura H,Tachibana M,Lorincz MC,Shinkai Y.Proviral silencing in embryonic stem cells requires the histone methyltransferase ESET.Nature,2010,464(7290):927-931.
����[21] Stadtfeld M,Maherali N,Breault DT,Hochedlinger K.Defining molecular cornerstones during fibroblast to i PS cell reprogramming in mouse.Cell Stem Cell,2008,2(3):230-240.
����[22] Sridharan R,Tchieu J,Mason MJ,Yachechko R,Kuoy E,Horvath S,Zhou Q,Plath K.Role of the murine reprogramming factors in the induction of pluripotency.Cell,2009,136(2):364-377.
����[23] Okita K,Nakagawa M,Hyenjong H,Ichisaka T,Yamanaka S.Generation of mouse induced pluripotent stem cells without viral vectors.Science,2008,322(5903):949-953.
����[24] Lee CS,Bishop ES,Zhang RY,Yu XY,Farina EM,Yan SJ,Zhao C,Zheng ZY,Shu Y,Wu XY,Lei JY,Li YS,Zhang WW,Yang C,Wu K,Wu Y,Ho S,Athiviraham A,Lee MJ,Wolf JM,Reid RR,He TC.Adenovirusmediated gene delivery:potential applications for gene and cell-based therapies in the new era of personalized medicine.Genes Dis,2017,4(2):43-63.
����[25] Somers A,Jean JC,Sommer CA,Omari A,Ford CC,Mills JA,Ying L,Sommer AG,Jean JM,Smith BW,Lafyatis R,Demierre MF,Weiss DJ,French DL,Gadue P,Murphy GJ,Mostoslavsky G,Kotton DN.Generation of transgene-free lung disease-specific human induced pluripotent stem cells using a single excisable lentiviral stem cell cassette.Stem Cells,2010,28(10):1728-1740.
����[26] Maherali N,Ahfeldt T,Rigamonti A,Utikal J,Cowan C,Hochedlinger K.A high-efficiency system for the generation and study of human induced pluripotent stem cells.Cell Stem Cell,2008,3(3):340-345.
����[27] Abbar AA,Nordin N,Ngai SC,Abdullah S.Production of lentiviral vector with polycistronic transcripts for reprogramming of mouse fibroblast cells.Pertanika JSci Technol,2018,26(2):627-640.
����[28] Kaji K,Norrby K,Paca A,Mileikovsky M,Mohseni P,Woltjen K.Virus-free induction of pluripotency and subsequent excision of reprogramming factors.Nature,2009,458(7239):771-775.
����[29] Soldner F,Hockemeyer D,Beard C,Gao Q,Bell GW,Cook EG,Hargus G,Blak A,Cooper O,Mitalipova M,Isacson O,Jaenisch R.Parkinson's disease patientderived induced pluripotent stem cells free of viral reprogramming factors.Cell,2009,136(5):964-977.
����[30] Wernig M,Lengner CJ,Hanna J,Lodato MA,Steine E,Foreman R,Staerk J,Markoulaki S,Jaenisch R.A druginducible transgenic system for direct reprogramming of multiple somatic cell types.Nat Biotechnol,2008,26(8):916-924.
����[31] Zhang X,De Los Angeles A,Zhang J.The art of human induced pluripotent stem cells:the past,the present and the future.Open Stem Cell J,2010,2:2-7.
����[32] Li J,Song W,Pan GJ,Zhou J.Advances in understanding the cell types and approaches used for generating induced pluripotent stem cells.J Hematol Oncol,2014,7:50.
����[33] Yamanaka S.A fresh look at i PS cells.Cell,2009,137(1):13-17.
����[34] Qian QJ,Che JQ,Ye LP,Zhong BX.The improvement and application of piggy Bac transposon system in mammals.Hereditas(Beijing),2014,36(10):965-973.Ǯ���,����ٻ,Ҷ¶��,�Ӳ���.piggy Bacת��ϵͳ�Ĺ��ܸĽ����ڲ��鶯���е�Ӧ��.�Ŵ�,2014,36(10):965-973.
����[35] Woltjen K,Michael IP,Mohseni P,Desai R,Mileikovsky M,H?m?l?inen R,Cowling R,Wang W,Liu PT,Gertsenstein M,Kaji K,Sung HK,Nagy A.Piggy Bac transposition reprograms fibroblasts to induced pluripotent stem cells.Nature,2009,458(7239):766-770.
����[36] Medvedev SP,Shevchenko AI,Zakian SM.Induced pluripotent stem cells:problems and advantages when applying them in regenerative medicine.Acta Nature,2010,2(2):18-27.
����[37] Davis RP,Nemes C,Varga E,Freund C,Kosmidis G,Gkatzis K,de Jong D,Szuhai K,Dinnyés A,Mummery CL.Generation of induced pluripotent stem cells from human foetal fibroblasts using the Sleeping Beauty transposon gene delivery system.Differentiation,2013,86(1-2):30-37.
����[38] Stadtfeld M,Nagaya M,Utikal J,Weir G,Hochedlinger K.Induced pluripotent stem cells generated without viral integration.Science,2008,322(5903):945-949.
����[39] Macarthur CC,Fontes A,Ravinder N,Kuninger D,Kaur J,Bailey M,Taliana A,Vemuri MC,Lieu PT.Generation of human-induced pluripotent stem cells by a nonintegrating RNA Sendai virus vector in feeder-free or xeno-free conditions.Stem Cells Int,2012,2012:1-9.
����[40] Kawagoe S,Higuchi T,Otaka M,Shimada Y,Kobayashi H,Ida H,Ohashi T,Okano HJ,Nakanishi M,Eto Y.Morphological features of i PS cells generated from Fabry disease skin fibroblasts using Sendai virus vector(Se Vdp).Mol Genet Metab,2013,109(4):386-389.
����[41] Yonemitsu Y,Kitson C,Ferrari S,Farley R,Griesenbach U,Judd D,Steel R,Scheid P,Zhu J,Jeffery PK,Kato A,Hasan MK,Nagai Y,Masaki I,Fukumura M,Hasegawa M,Geddes DM,Alton EW.Efficient gene transfer to airway epithelium using recombinant Sendai virus.Nat Biotechnol,2000,18(9):970-973.
����[42] Takeda A,Igarashi H,Kawada M,Tsukamoto T,Yamamoto H,Inoue M,Iida A,Shu T,Hasegawa M,Matano T.Evaluation of the immunogenicity of replication-competent V-knocked-out and replicationdefective F-deleted Sendai virus vector-based vaccines in macaques.Vaccine,2008,26(52):6839-6843.
����[43] Nakanishi M,Otsu M.Development of Sendai virus vectors and their potential applications in gene therapy and regenerative medicine.Curr Gene Ther,2012,12(5):410-416.
����[44] Yu JY,Hu KJ,Smuga-Otto K,Tian SL,Stewart R,Slukvin II,Thomson JA.Human induced pluripotent stem cells free of vector and transgene sequences.Science,2009,324(5928):797-801.
����[45] Jia FJ,Wilson KD,Sun N,Gupta DM,Huang M,Li ZJ,Panetta NJ,Chen ZY,Robbins RC,Kay MA,Longaker MT,Wu JC.A nonviral minicircle vector for deriving human i PS cells.Nat Methods,2010,7(3):197-199.
����[46] Zhou WB,Freed CR.Adenoviral gene delivery can reprogram human fibroblasts to induced pluripotent stem cells.Stem Cells,2009,27(11):2667-2674.
����[47] Cho HJ,Lee CS,Kwon YW,Paek JS,Lee SH,Hur J,Lee EJ,Roh TY,Chu IS,Leem SH,Kim Y,Kang HJ,Park YB,Kim HS.Induction of pluripotent stem cells from adult somatic cells by protein-based reprogramming without genetic manipulation.Blood,2010,116(3):386-395.
����[48] Kim D,Kim CH,Moon JI,Chung YG,Chang MY,Han BS,Ko S,Yang E,Cha KY,Lanza R,Kim KS.Generation of human induced pluripotent stem cells by direct delivery of reprogramming proteins.Cell Stem Cell,2009,4(6):472-476.
����[49] Mandal PK,Rossi DJ.Reprogramming human fibroblasts to pluripotency using modified m RNA.Nat Protoc,2013,8(3):568-582.
����[50] Ong SG,Lee WH,Kodo K,Wu JC.Micro RNA-mediated regulation of differentiation and transdifferentiation in stem cells.Adv Drug Deliv Rev,2015,88:3-15.
����[51] Huangfu D,Osafune K,Maehr R,Guo W,Eijkelenboom A,Chen SB,Muhlestein W,Melton DA.Induction of pluripotent stem cells from primary human fibroblasts with only Oct4 and Sox2.Nat Biotechnol,2008,26(11):1269-1275.
����[52] Huangfu D,Maehr R,Guo WJ,Eijkelenboom A,Snitow M,Chen AE,Melton DA.Induction of pluripotent stem cells by defined factors is greatly improved by smallmolecule compounds.Nat Biotechnol,2008,26(7):795-797.
����[53] Mali P,Chou BK,Yen J,Ye ZH,Zou JZ,Dowey S,Brodsky RA,Ohm JE,Yu W,Baylin SB,Yusa K,Bradley A,Meyers DJ,Mukherjee C,Cole PA,Cheng LZ.Butyrate greatly enhances derivation of human induced pluripotent stem cells by promoting epigenetic remodeling and the expression of pluripotency-associated genes.Stem Cells,2010,28(4):713-720.
����[54] Yoshida Y,Takahashi K,Okita K,Ichisaka T,Yamanaka S.Hypoxia enhances the generation of induced pluripotent stem cells.Cell Stem Cell,2009,5(3):237-241.
����[55] Hong H,Takahashi K,Ichisaka T,Aoi T,Kanagawa O,Nakagawa M,Okita K,Yamanaka S.Suppression of induced pluripotent stem cell generation by the p53-p21pathway.Nature,2009,460(7259):1132-1135.
����[56] Rais Y,Zviran A,Geula S,Gafni O,Chomsky E,Viukov S,Mansour AA,Caspi I,Krupalnik V,Zerbib M,Maza I,Mor N,Baran D,Weinberger L,Jaitin DA,Lara-Astiaso D,Blecher-Gonen R,Shipony Z,Mukamel Z,Hagai T,Gilad S,Amann-Zalcenstein D,Tanay A,Amit I,Novershtern N,Hanna JH.Deterministic direct reprogramming of somatic cells to pluripotency.Nature,2013,502(7469):65-70
����[57] Maekawa M,Yamaguchi K,Nakamura T,Shibukawa R,Kodanaka I,Ichisaka T,Kawamura Y,Mochizuki H,Goshima N,Yamanaka S.Direct reprogramming of somatic cells is promoted by maternal transcription factor Glis1.Nature,2011,474(7350):225-229.
����[58] Kunitomi A,Yuasa S,Sugiyama F,Saito Y,Seki T,Kusumoto D,Kashimura S,Takei M,Tohyama S,Hashimoto H,Egashira T,Tanimoto Y,Mizuno S,Tanaka S,Okuno H,Yamazawa K,Watanabe H,Oda M,Kaneda R,Matsuzaki Y,Nagai T,Okano H,Yagami KI,Tanaka M,Fukuda K.H1foo has a pivotal role in qualifying induced pluripotent stem cells.Stem Cell Reports,2016,6(6):825-833
����[59] Maherali N,Hochedlinger K.Tgfbeta signal inhibition cooperates in the induction of i PSCs and replaces Sox2and c Myc.Curr Biol,2009,19(20):1718-1723.
����[60] Rajasingh J,Thangavel J,Siddiqui MR,Gomes I,Gao XP,Kishore R,Malik AB.Improvement of cardiac function in mouse myocardial infarction after transplantation of epigenetically-modified bone marrow progenitor cells.PLo S One,2011,6(7):e22550.
����[61] Anokye-Danso F,Trivedi CM,Juhr D,Gupta M,Cui Z,Tian Y,Zhang YZ,Yang WL,Gruber PJ,Epstein JA,Morrisey EE.Highly efficient mi RNA-mediated reprogramming of mouse and human somatic cells to pluripotency.Cell Stem Cell,2011,8(4):376-388.
����[62] Hou PP,Li YQ,Zhang X,Liu C,Guan JY,Li HG,Zhao T,Ye JQ,Yang WF,Liu K,Ge J,Xu J,Zhang Q,Zhao Y,Deng HK.Pluripotent stem cells induced from mouse somatic cells by small-molecule compounds.Science,2013,341(6146):651-654.
����[63] Hockemeyer D,Jaenisch R.Induced pluripotent stem cells meet genome editing.Cell Stem Cell,2016,18(5):573-586.
����[64] Hockemeyer D,Soldner F,Beard C,Gao Q,Mitalipova M,De Kelver RC,Katibah GE,Amora R,Boydston EA,Zeitler B,Meng XD,Miller JC,Zhang L,Rebar EJ,Gregory PD,Urnov FD,Jaenisch R.Efficient targeting of expressed and silent genes in human ESCs and i PSCs using zinc-finger nucleases.Nat Biotechnol,2009,27(9):851-857.
����[65] Zou JZ,Maeder ML,Mali P,Pruett-Miller SM,Thibodeau-Beganny S,Chou BK,Chen GB,Ye ZH,Park IH,Daley GQ,Porteus MH,Joung JK,Cheng LZ.Gene targeting of a disease-related gene in human induced pluripotent stem and embryonic stem cells.Cell Stem Cell,2009,5(1):97-110.
����[66] Christian M,Cermak T,Doyle EL,Schmidt C,Zhang F,Hummel A,Bogdanove AJ,Voytas DF.Targeting DNAdouble-strand breaks with TAL effector nucleases.Genetics,2010,186(2):757-761.
����[67] Hockemeyer D,Wang HY,Kiani S,Lai CS,Gao Q,Cassady JP,Cost GJ,Zhang L,Santiago Y,Miller JC,Zeitler B,Cherone JM,Meng XD,Hinkley SJ,Rebar EJ,Gregory PD,Urnov FD,Jaenisch R.Genetic engineering of human pluripotent cells using TALEnucleases.Nat Biotechnol,2011,29(8):731-734.
����[68] Cong L,Ran FA,Cox D,Lin SL,Barretto R,Habib N,Hsu PD,Wu XB,Jiang WY,Marraffini LA,Zhang F.Multiplex genome engineering using CRISPR/Cas systems.Science,2013,339(6121):819-823.
����[69] Fu YF,Foden JA,Khayter C,Maeder ML,Reyon D,Joung JK,Sander JD.High-frequency off-target mutagenesis induced by CRISPR-Cas nucleases in human cells.Nat Biotechnol,2013,31(9):822-826.
����[70] Smith C,Gore A,Yan W,Abalde-Atristain L,Li Z,He CX,Wang Y,Brodsky RA,Zhang K,Cheng LZ,Ye ZH.Whole-genome sequencing analysis reveals high specificity of CRISPR/Cas9 and TALEN-based genome editing in human i PSCs.Cell Stem Cell,2014,15(1):12-13.
����[71] Veres A,Gosis BS,Ding QR,Collins R,Ragavendran A,Brand H,Erdin S,Cowan CA,Talkowski ME,Musunuru K.Low incidence of off-target mutations in inpidual CRISPR-Cas9 and TALEN targeted human stem cell clones detected by whole-genome sequencing.Cell Stem Cell,2014,15(1):27-30.
����[72] Ran FA,Hsu PD,Lin CY,Gootenberg JS,Konermann S,Trevino AE,Scott DA,Inoue A,Matoba S,Zhang Y,Zhang F.Double nicking by RNA-guided CRISPR Cas9for enhanced genome editing specificity.Cell,2013,154(6):1380-1389.
����[73] Slaymaker IM,Gao LY,Zetsche B,Scott DA,Yan WX,Zhang F.Rationally engineered Cas9 nucleases with improved specificity.Science,2016,351(6268):84-88.
����[74] Chen BH,Gilbert LA,Cimini BA,chnitzbauer J,Zhang W,Li GW,Park J,Blackburn EH,Weissman JS,Qi LS,Huang B.Dynamic imaging of genomic loci in living human cells by an optimized CRISPR/Cas system.Cell,2013,155(7):1479-1491.
����[75] Paquet D,Kwart D,Chen A,Sproul A,Jacob S,Teo S,Olsen KM,Gregg A,Noggle S,Tessier-Lavigne M.Efficient introduction of specific homozygous and heterozygous mutations using CRISPR/Cas9.Nature,2016,533(7601):125-129.
����[76] Komor AC,Kim YB,Packer MS,Zuris JA,Liu DR.Programmable editing of a target base in genomic DNAwithout double-stranded DNA cleavage.Nature,2016,533(7603):420-424.
����[77] Ebert AD,Yu JY,Rose FF Jr,Mattis VB,Lorson CL,Thomson JA,Svendsen CN.Induced pluripotent stem cells from a spinal muscular atrophy patient.Nature,2009,457(7227):277-280.
����[78] Munsat TL,Davies KE.International SMA consortium meeting.Neuromuscul Disord,1992,2(5-6):423-428.
����[79] Yang XL,Pabon L,Murry CE.Engineering adolescence:maturation of human pluripotent stem cell-derived cardiomyocytes.Circ Res,2014,114(3):511-523.
����[80] Funakoshi S,Miki K,Takaki T,Okubo C,Hatani T,Chonabayashi K,Nishikawa M,Takei I,Oishi A,Narita M,Hoshijima M,Kimura T,Yamanaka S,Yoshida Y.Enhanced engraftment,proliferation,and therapeutic potential in heart using optimized human i PSC-derived cardiomyocytes.Sci Rep,2016,6:19111.
����[81] Yang XL,Rodriguez M,Pabon L,Fischer KA,Reinecke H,Regnier M,Sniadecki NJ,Ruohola-Baker H,Murry CE.Tri-iodo-l-thyronine promotes the maturation of human cardiomyocytes-derived from induced pluripotent stem cells.J Mol Cell Cardiol,2014,72:296-304.
����[82] Kamakura T,Makiyama T,Sasaki K,Yoshida Y,Wuriyanghai Y,Chen JR,Hattori T,Ohno S,Kita T,Horie M,Yamanaka S,Kimura T.Ultrastructural maturation of human-induced pluripotent stem cell-derived cardiomyocytes in a long-term culture.Circ J,2013,77(5):1307-1314.
����[83] Nunes SS,Miklas JW,Liu J,Aschar-Sobbi R,Xiao Y,Zhang B,Jiang J,MasséS,Gagliardi M,Hsieh A,Thavandiran N,Laflamme MA,Nanthakumar K,Gross GJ,Backx PH,Keller G,Radisic M.Biowire:a platform for maturation of human pluripotent stem cell-derived cardiomyocytes.Nat Methods,2013,10(8):781-787.
����[84] Pearson BL,Simon JM,Mc Coy ES,Salazar G,Fragola G,Zylka MJ.Identification of chemicals that mimic transcriptional changes associated with autism,brain aging and neurodegeneration.Nat Commun,2016,7:11173.
����[85] Miller JD,Ganat YM,Kishinevsky S,Bowman RL,Liu B,Tu EY,Mandal PK,Vera E,Shim JW,Kriks S,Taldone T,Fusaki N,Tomishima MJ,Krainc D,Milner TA,Rossi DJ,Studer L.Human i PSC-based modeling of late-onset disease via progerin-induced aging.Cell Stem Cell,2013,13(6):691-705.
����[86] Kondo T,Asai M,Tsukita K,Kutoku Y,Ohsawa Y,Sunada Y,Imamura K,Egawa N,Yahata N,Okita K,Takahashi K,Asaka I,Aoi T,Watanabe A,Watanabe K,Kadoya C,Nakano R,Watanabe D,Maruyama K,Hori O,Hibino S,Choshi T,Nakahata T,Hioki H,Kaneko T,Naitoh M,Yoshikawa K,Yamawaki S,Suzuki S,Hata R,Ueno S,Seki T,Kobayashi K,Toda T,Murakami K,Irie K,Klein WL,Mori H,Asada T,Takahashi R,Iwata N,Yamanaka S,Inoue H.Modeling Alzheimer's disease with i PSCs reveals stress phenotypes associated with intracellular Aβand differential drug responsiveness.Cell Stem Cell,2013,12(4):487-496.
����[87] Soldner F,Stelzer Y,Shivalila CS,Abraham BJ,Latourelle JC,Barrasa MI,Goldmann J,Myers RH,Young RA,Jaenisch R.Parkinson-associated risk variant in distal enhancer ofα-synuclein modulates target gene expression.Nature,2016,533(7601):95-99.
����[88] Gandre-Babbe S,Paluru P,Aribeana C,Chou ST,Bresolin S,Lu L,Sullivan SK,Tasian SK,Weng JL,Favre H,Choi JK,French DL,Loh ML,Weiss MJ.Patient-derived induced pluripotent stem cells recapitulate hematopoietic abnormalities of juvenile myelomonocytic leukemia.Blood,2013,121(24):4925-4929.
����[89] Malkin D.p53 and the Li-Fraumeni syndrome.Cancer Genet Cytogenet,1993,66(2):83-92.
����[90] Lee DF,Su J,Kim HS,Chang B,Papatsenko D,Zhao RY,Yuan Y,Gingold J,Xia WY,Darr H,Mirzayans R,Hung MC,Schaniel C,Lemischka IR.Modeling familial cancer with induced pluripotent stem cells.Cell,2015,161(2):240-254.
����[91] Mc Cauley HA,Wells JM.Pluripotent stem cell-derived organoids:using principles of developmental biology to grow human tissues in a dish.Development,2017,144(6):958-962.
����[92] Dutta D,Heo I,Clevers H.Disease modeling in stem cell-derived 3D organoid systems.Trends Mol Med,2017,23(5):393-410.
����[93] Lee G,Papapetrou EP,Kim H,Chambers SM,Tomishima MJ,Fasano CA,Ganat YM,Menon J,Shimizu F,Viale A,Tabar V,Sadelain M,Studer L.Modelling pathogenesis and treatment of familial dysautonomia using patient-specific i PSCs.Nature,2009,461(7262):402-406.
����[94] Lancaster MA,Renner M,Martin CA,Wenzel D,Bicknell LS,Hurles ME,Homfray T,Penninger JM,Jackson AP,Knoblich JA.Cerebral organoids model human brain development and microcephaly.Nature,2013,501(7467):373-379.
����[95] Quadrato G,Brown J,Arlotta P.The promises and challenges of human brain organoids as models of neuropsychiatric disease.Nat Med,2016,22(11):1220-1228.
����[96] Sloan SA,Darmanis S,Huber N,Khan TA,Birey F,Caneda C,Reimer R,Quake SR,Barres BA,Pa?ca SP.Human astrocyte maturation captured in 3D cerebral cortical spheroids derived from pluripotent stem cells.Neuron,2017,95(4):779-790.
����[97] Gabriel E,Wason A,Ramani A,Gooi LM,Keller P,Pozniakovsky A,Poser I,Noack F,Telugu NS,Calegari F,?ari?T,Hescheler J,Hyman AA,Gottardo M,Callaini G,Alkuraya FS,Gopalakrishnan J.CPAPpromotes timely cilium disassembly to maintain neural progenitor pool.EMBO J,2016,35(8):803-819.
����[98] Bian S,Repic M,Guo ZM,Kavirayani A,Burkard T,Bagley JA,Krauditsch C,Knoblich JA.Genetically engineered cerebral organoids model brain tumor formation.Nat Methods,2018,15(8):631-639.
����[99] Rashid ST,Corbineau S,Hannan N,Marciniak SJ,Miranda E,Alexander G,Huang-Doran I,Griffin J,Ahrlund-Richter L,Skepper J,Semple R,Weber A,Lomas DA,Vallier L.Modeling inherited metabolic disorders of the liver using human induced pluripotent stem cells.J Clin Invest,2010,120(9):3127-3136.
����[100] Ogawa M,Ogawa S,Bear CE,Ahmadi S,Chin S,Li B,Grompe M,Keller G,Kamath BM,Ghanekar A.Directed differentiation of cholangiocytes from human pluripotent stem cells.Nat Biotechnol,2015,33(8):853-861.
����[101] Sampaziotis F,de Brito MC,Madrigal P,Bertero A,Saeb-Parsy K,Soares FAC,Schrumpf E,Melum E,Karlsen TH,Bradley JA,Gelson WT,Davies S,Baker A,Kaser A,Alexander GJ,Hannan NRF,Vallier L.Cholangiocytes derived from human induced pluripotent stem cells for disease modeling and drug validation.Nat Biotechnol,2015,33(8):845-852.
����[102] Voges HK,Mills RJ,Elliott DA,Parton RG,Porrello ER,Hudson JE.Development of a human cardiac organoid injury model reveals innate regenerative potential.Development,2017,144(6):1118-1127.
����[103] Pagliuca FW,Millman JR,Gürtler M,Segel M,Van Dervort A,Ryu JH,Peterson QP,Greiner D,Melton DA.Generation of functional human pancreaticβcells in vitro.Cell,2014,159(2):428-439
����[104] Hoang P,Wang J,Conklin BR,Healy KE,Ma Z.Generation of spatial-patterned early-developing cardiac organoids using human pluripotent stem cells.Nat Protoc,2018,13(4):723-737.
����[105] Abilez OJ,Tzatzalos E,Yang HX,Zhao MT,Jung G,Z?llner AM,Tiburcy M,Riegler J,Matsa E,Shukla P,Zhuge Y,Chour T,Chen VC,Burridge PW,Karakikes I,Kuhl E,Bernstein D,Couture LA,Gold JD,Zimmermann WH,Wu JC.Passive stretch induces structural and functional maturation of engineered heart muscle as predicted by computational modeling.Stem Cells,2018,36(2):265-277.
����[106] Musah S,Mammoto A,Ferrante TC,Jeanty SSF,Hirano-Kobayashi M,Mammoto T,Roberts K,Chung S,Novak R,Ingram M,Fatanat-Didar T,Koshy S,Weaver JC,Church GM,Ingber DE.Mature induced-pluripotentstem-cell-derived human podocytes reconstitute kidney glomerular-capillary-wall function on a chip.Nat Biomed Eng,2017,1:0069.
����[107] Brown JA,Pensabene V,Markov DA,Allwardt V,Neely MD,Shi MJ,Britt CM,Hoilett OS,Yang Q,Brewer BM,Samson PC,Mc Cawley LJ,May JM,Webb DJ,Li DY,Bowman AB,Reiserer RS,Wikswo JP.Recreating blood-brain barrier physiology and structure on chip:a novel neurovascular microfluidic bioreactor.Biomicrofluidics,2015,9(5):054124.
����[108] Mc Cauley KB,Hawkins F,Serra M,Thomas DC,Jacob A,Kotton DN.Efficient derivation of functional human airway epithelium from pluripotent stem cells via temporal regulation of Wnt signaling.Cell Stem Cell,2017,20(6):844-857.
����[109] Jacob A,Morley M,Hawkins F,Mc Cauley KB,Jean JC,Heins H,Na CL,Weaver TE,Vedaie M,Hurley K,Hinds A,Russo SJ,Kook S,Zacharias W,Ochs M,Traber K,Quinton LJ,Crane A,Davis BR,White FV,Wambach J,Whitsett JA,Cole FS,Morrisey EE,Guttentag SH,Beers MF,Kotton DN.Differentiation of human pluripotent stem cells into functional lung alveolar epithelial cells.Cell Stem Cell,2017,21(4):472-488.
����[110] Liu C,Oikonomopoulos A,Sayed N,Wu JC.Modeling human diseases with induced pluripotent stem cells:from 2D to 3D and beyond.Development,2018,145(5):dev156166.
����[111] Sturgeon CM,Ditadi A,Awong G,Kennedy M,Keller G.Wnt signaling controls the specification of definitive and primitive hematopoiesis from human pluripotent stem cells.Nat Biotechnol,2014,32(6):554-561.
����[112] Wang LS,Li L,Menendez P,Cerdan C,Bhatia M.Human embryonic stem cells maintained in the absence of mouse embryonic fibroblasts or conditioned media are capable of hematopoietic development.Blood,2005,105(12):4598-4603.
����[113] Barker RA,Parmar M,Studer L,Takahashi J.Human trials of stem cell-derived dopamine neurons for Parkinson’s disease:dawn of a new era.Cell Stem Cell,2017,21(5):569-573.
����[114] Espuny-Camacho I,Michelsen KA,Gall D,Linaro D,Hasche A,Bonnefont J,Bali C,Orduz D,Bilheu A,Herpoel A,Lambert N,Gaspard N,Péron S,Schiffmann SN,Giugliano M,Gaillard A,Vanderhaeghen P.Pyramidal neurons derived from human pluripotent stem cells integrate efficiently into mouse brain circuits in vivo.Neuron,2013,77(3):440-456.
����[115] Yuan T,Liao W,Feng NH,Lou YL,Niu X,Zhang AJ,Wang Y,Deng ZF.Human induced pluripotent stem cell-derived neural stem cells survive,migrate,differentiate,and improve neurologic function in a rat model of middle cerebral artery occlusion.Stem Cell Res Ther,2013,4(3):73.
����[116] Sundberg M,Bogetofte H,Lawson T,Jansson J,Smith G,Astradsson A,Moore M,Osborn T,Cooper O,Spealman R,Hallett P,Isacson O.Improved cell therapy protocols for Parkinson’s disease based on differentiation efficiency and safety of h ESC-,hi PSC-,and non-human primate i PSC-derived dopaminergic neurons.Stem Cells,2013,31(8):1548-1562.
����[117] Donegan JJ,Lodge DJ.Cell-based therapies for the treatment of schizophrenia.Brain Res,2017,1655:262-269.
����[118] Mansour AA,Gon?alves JT,Bloyd CW,Li H,Fernandes S,Quang D,Johnston S,Parylak SL,Jin X,Gage FH.An in vivo model of functional and vascularized human brain organoids.Nat Biotechnol,2018,36(5):432-441.
����[119] Takebe T,Sekine K,Enomura M,Koike H,Kimura M,Ogaeri T,Zhang RR,Ueno Y,Zheng YW,Koike N,Aoyama S,Adachi Y,Taniguchi H.Vascularized and functional human liver from an i PSC-derived organ bud transplant.Nature,2013,499(7459):481-484.
����[120] Yang JY,Wang Y,Zhou T,Wong LY,Tian XY,Hong XY,Lai WH,Au KW,Wei R,Liu YQ,Cheng LH,Liang GC,Huang ZJ,Fan WX,Zhao P,Wang XW,Iba?ez DP,Luo ZW,Li YY,Zhong XF,Chen SH,Wang DY,Li L,Lai LX,Qin BM,Bao XC,Hutchins AP,Siu CW,Huang Y,Esteban MA,Tse HF.Generation of human liver chimeric mice with hepatocytes from familial hypercholesterolemia induced pluripotent stem cells.Stem Cell Reports,2017,8(3):605-618
����[121] Chao MP,Gentles AJ,Chatterjee S,Lan F,Reinisch A,Corces MR,Xavy S,Shen J,Haag D,Chanda S,Sinha R,Morganti RM,Nishimura T,Ameen M,Wu HD,Wernig M,Wu JC,Majeti R.Human AML-i PSCs reacquire leukemic properties after differentiation and model clonal variation of disease.Cell Stem Cell,2017,20(3):329-344.
����[122] Stricker SH,Feber A,Engstr?m PG,Carén H,Kurian KM,Takashima Y,Watts C,Way M,Dirks P,Bertone P,Smith A,Beck S,Pollard SM.Widespread resetting of DNA methylation in glioblastoma-initiating cells suppresses malignant cellular behavior in a lineagedependent manner.Genes Dev,2013,27(6):654-669.
����[123] Lee DF,Su J,Kim HS,Chang B,Papatsenko D,Zhao RY,Yuan Y,Gingold J,Xia WY,Darr H,Mirzayans R,Hung MC,Schaniel C,Lemischka IR.Modeling familial cancer with induced pluripotent stem cells.Cell,2015,161(2):240-254.
����[124] Kiskinis E,Eggan K.Progress toward the clinical application of patient-specific pluripotent stem cells.JClin Invest,2010,120(1):51-59.
����[125] Lee AS,Tang C,Rao MS,Weissman IL,Wu JC.Tumorigenicity as a clinical hurdle for pluripotent stem cell therapies.Nat Med,2013,19(8):998-1004.
����[126] Lund RJ,N?rv?E,Lahesmaa R.Genetic and epigenetic stability of human pluripotent stem cells.Nat Rev Genet,2012,13(10):732-744.
����[127] Hotta A,Yamanaka S.From genomics to gene therapy:induced pluripotent stem cells meet genome editing.Annu Rev Genet,2015,49:47-70.
����[128] Okita K,Matsumura Y,Sato Y,Okada A,Morizane A,Okamoto S,Hong H,Nakagawa M,Tanabe K,Tezuka K,Shibata T,Kunisada T,Takahashi M,Takahashi J,Saji H,Yamanaka S.A more efficient method to generate integration-free human i PS cells.Nat Methods,2011,8(5):409-412.
����[129] Kim D,Lee DR,Kim HS,Yoo J,Jung SJ,Lim BY,Jang J,Kang HC,You S,Hwang DY,Leem JW,Nam TS,Cho SR,Kim DW.Highly pure and expandable PSA-NCAM-positive neural precursors from human ESC and i PSC-derived neural rosettes.PLo S One,2012,7(7):e39715.
����[130] Doi D,Samata B,Katsukawa M,Kikuchi T,Morizane A,Ono Y,Sekiguchi K,Nakagawa M,Parmar M,Takahashi J.Isolation of human induced pluripotent stem cell-derived dopaminergic progenitors by cell sorting for successful transplantation.Stem Cell Reports,2014,2(3):337-350.
����[131] Finkbeiner SR,Hill DR,Altheim CH,Dedhia PH,Taylor MJ,Tsai Y,Chin AM,Mahe MM,Watson CL,Freeman JJ,Nattiv R,Thomson M,Klein OD,Shroyer NF,Helmrath MA,Teitelbaum DH,Dempsey PJ,Spence JR.Transcriptome-wide analysis reveals hallmarks of human intestine development and maturation in vitro and in vivo.Stem Cell Reports,2015,4(6):1140-1155.
����[132] Mandai M,Watanabe A,Kurimoto Y,Hirami Y,Morinaga C,Daimon T,Fujihara M,Akimaru H,Sakai N,Shibata Y,Terada M,Nomiya Y,Tanishima S,Nakamura M,Kamao H,Sugita S,Onishi A,Ito T,Fujita K,Kawamata S,Go MJ,Shinohara C,Hata KI,Sawada M,Yamamoto M,Ohta S,Ohara Y,Yoshida K,Kuwahara J,Kitano Y,Amano N,Umekage M,Kitaoka F,Tanaka A,Okada C,Takasu N,Ogawa S,Yamanaka S,Takahashi M.Autologous induced stem-cell-derived retinal cells for macular degeneration.N Engl J Med,2017,376(11):1038-1046
����[133] Kuriyan AE,Albini TA,Townsend JH,Rodriguez M,Pandya HK,Leonard RE,Parrott MB,Rosenfeld PJ,Flynn HW Jr,Goldberg JL.Vision loss after intravitreal injection of autologous“Stem Cells”for AMD.N Engl JMed,2017,376(11):1047-1053.
����[134] Cell Stem Cell Editorial Team.10 Questions:Clinical Outlook for i PSCs.Cell Stem Cell,2016,18(2):170-173.
����[135] Themeli M,Kloss CC,Ciriello G,Fedorov VD,Perna F,Gonen M,Sadelain M.Generation of tumor-targeted human T lymphocytes from induced pluripotent stem cells for cancer therapy.Nat Biotechnol,2013,31(10):928-933.
����[136] Nianias A,Themeli M.Induced pluripotent stem cell(i PSC)-derived lymphocytes for adoptive cell immunotherapy:recent advances and challenges.Curr Hematol Malig Rep,2019,14(4):261-268.
����[137] Minagawa A,Yoshikawa T,Yasukawa M,Hotta A,Kunitomo M,Iriguchi S,Takiguchi M,Kassai Y,Imai E,Yasui Y,Kawai Y,Zhang R,Uemura Y,Miyoshi H,Nakanishi M,Watanabe A,Hayashi A,Kawana K,Fujii T,Nakatsura T,Kaneko S.Enhancing T cell receptor stability in rejuvenated i PSC-derived T cells improves their use in cancer immunotherapy.Cell Stem Cell,2018,23(6):850-858.e4.
����[138] News and events.Ci RA,Kyoto University.2019.Available at:www.cira.kyoto-u.ac.jp/e/pressrelease/news/190716-150000.html (accessed September25,2019).
����[139] Morvan MG,Lanier LL.NK cells and cancer:you can teach innate cells new tricks.Nat Rev Cancer,2016,16(1):7-19.
����[140] Lupo KB,Matosevic S.Natural killer cells as allogeneic effectors in adoptive cancer immunotherapy.Cancers(Basel),2019,11(6):769.
����[141] Chen S,Sun H,Miao K,Deng CX.CRISPR-Cas9:from genome editing to cancer research.Int J Biol Sci,2016,12(12):1427-1436.
����[142] Li Y,Hermanson DL,Moriarity BS,Kaufman DS.Human i PSC-derived natural killer cells engineered with chimeric antigen receptors enhance anti-tumor activity.Cell Stem Cell,2018,23(2):181-192.e5.
����[143] Lo Sardo V,Ferguson W,Erikson GA,Topol EJ,Baldwin KK,Torkamani A.Influence of donor age on induced pluripotent stem cells.Nature Biotechnology,2017,35(1):69-74.
����[144] Hanahan D,Weinberg RA.Hallmarks of cancer:the next generation.Cell,2011,144(5):646-674.





