2.1调节昼夜节律
早在1970年,LiptonG.R.对蟑螂额神经节电生理学研究发现,额神经节在没有任何信号输入的情况下能自发放电,认为它可能是一个具有自律性的神经结构。后来的研究发现蟑螂每日的行为、自发放电的时期及额神经节内5-HT的含量,都存在周期节律性的变化[4,16].并认为5-HT与调节昆虫的昼夜节律有关[17].近年,SaritA.进一步研究发现,sg-inx1、sg-inx2、sg-inx3、sg-inx4基因在额神经节内表达缝隙连接蛋白[18-19],缝隙连接蛋白具有调节神经节律性的作用。因此,额神经节具有通过调节5-HT和缝隙连接蛋白来控制昼夜节律的功能。
2.2控制前肠的自律性运动
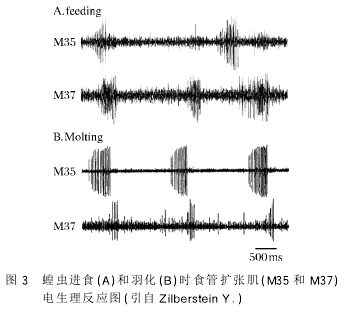
昆虫的消化道根 据其发生 的 来 源 和 机 能 不同,可分为前肠、中肠和后肠。前肠具有复杂的结构。蝗虫的前肠起始于口,经由咽喉、1个短而狭窄弯曲的食管、嗉囊,终止于 前 胃,与 中 肠 相 连。前肠的肌肉非常发达,共有2层,一层呈纵向排列,另一层呈环形排列[2,20].前肠的功能是磨碎食物,食物借助前肠的蠕动被消化。昆虫的进食和羽化行为都需要前肠肌肉的参与[21].所有支配前肠肌肉组织的神经都是来自于额神经节神经元。ZilbersteinY.等[22]应用电生理技术记录到了昆虫进食和羽化时前肠括约肌和食管扩张肌的节律性活动(图3)。去除昆虫的额神经节之后,昆虫的进食减少,羽化行为停止。羽化时,额神经节发出信号,使前肠扩张肌和括约肌收缩,导致昆虫吞咽行为。昆虫吞咽空气的行为使前肠充满空气,昆虫体内产生足够的压力使身体膨胀,裂开旧的外壳。因此,额神经节具有调控前肠肌肉运动进而调控昆虫的进食和羽化行为的功能。
2.3控制蛋白质的新陈代谢
Cedric G.等[23]发现当去除蝗虫的额神经节后,血液中的蛋白质水平迅速且永久性地降低;细胞质中的RNA浓度、血液氨基酸水平和中肠的蛋白酶含量也下降,而且通过喂食不能恢复。额神经节可能通过调节某种神经分泌物质的含量,来改变蛋白酶的量,进一步调控蛋白质的代谢。因此,额神经节对调控发育蝗虫的蛋白质新陈代谢具有重要的作用。
2.4调控心跳
昆虫的循环系统是开放式循环,包括背血管和血淋巴。背血管位于昆虫的背壁下方,是一条从头部到腹部的管状 结 构,分 为 心 脏 和 动 脉。心 脏 是 背 血 管 中 呈 连 续 膨 大 的 部 分,每 个 膨 大 的 部 分 称 为心室[24].
内脏神经是指分布于内脏、心血管和腺体的神经,属于交感神经系统。额神经节发出一条内脏神经,它在主动脉处分支为心前神经和心后神经[25].Kazuyuki U.等[25]通过对鳞翅目家蚕神经染色发现,心前神经支配背血管的前区,心前神经运动神经元的胞体位于额神经节。因此,背血管前区很可能由额神经节内的心前神经运动神经元支配,调控家蚕心跳动。
3结语
额神经节作为昆虫神经系统内的重要神经结构,多年来,许多研究者对额神经节的结构和功能进行了大量研究,虽然已经初步证明了额神经节在昆虫的进食、羽化、蛋白质代谢、昼夜节律调节等方面发挥重要作用,但由于额神经节多样性和复杂性及研究技术的限制,到现在为止,它的微细结构及各种功能的神经生物学机制尚未完全清楚,依然存在许多需要解决的问题。例如:从结构上看,需要搞清运动神经元和内分泌神经元的微细结构与分布,从功能上看,要阐明内分泌神经元的自律性调节机制等。为了探索额神经节的调控机制和神经传导机制,近年,Kanako M.等[26]应用cDNA减法杂交技术发现蚕额神经节基因(BmK5)表达神经肽(GSGTKVGGAGAATKVVTKSGS-NH2)和核定位蛋白。神经调节因子促使体外培养的额神经节输出运动信号,说明额神经节可能分泌神经调节因子发挥功能。这些发现,提示额神经节可能是一个内分泌器官,它通过分泌神经调节因子发挥作用,对其神经机制的进一步研究将是额神经节今后的研究方向和发展趋势。
参考文献:
[1]MILES C I,BOOKER R.The role of the frontal ganglion in foregut movements of the moth,Manduca sexta[J].JCompPhysiol A,1994,174(6):755-767.
[2]KENNETH U,GILLOTT C.Studies on the effects of the removal of the frontal ganglion in Locusta migratoria L[J].J ExpBiol,1967,46(1):13-25.
[3]BELL R A,BORG T K,ITTYCHERIAH P I.Neurosecretorycells in the frontal ganglion of the tobaccohornworm,Manduca sexta[J].J Insect Physiol,1974,20(4):669-675.
[4]PANDEY A,HABIBULLA M.Circadian rhythms of serotonin and the electrical activityof the frontal ganglion ofthe cockroach,Periplaneta americana[J].Cell Mol Life Sci,1982,38(8):946-948.
[5]OHANAA M,MOSERB T.Current and future imagingof the peripheral nervous system[J].Diagn Interv Imaging,2014,95(1):17-26.
[6]AYALI A.The insect frontal ganglion and stomatogastric pattern generator networks[J].Neurosignals,2004,13(1):20-36.
[7]COPINI P,KLAASSEN U,OUDEN J,et al.Precision of datinginsect outbreaks usingwood anatomy:the case ofAnoplophora in Japanese maple[J].Trees-Struct Funct,2014,28(1):103-113.
[8]MELTEM K,HATICE K.Investigation of the histological development of the frontal ganglion in Locusta migratoriaL.1758 (Orthoptera,Acrididae)[J].Turk J Zool,2012,36(2):241-248.
[9]PENZLIN H,STOLZNER W.Frontal ganglion and water balance in Periplaneta americana L[J].Cell Mol Life Sci,1971,27(4):390-391.
[10]NORMAN T D,JOHN G H.Neuroanatomyof the suckingpumpof the moth,Manduca se(Sphingidae,Lepidoptera)[J].Arthropod Struct Dev,2006,35(1):15-33.
[11]ITO K,SHINOMIYA K.A systematic nomenclature for the insect brain[J].Neuron,2014,81(4):755-765.
[12]GUNDEL M,PENZLIN H.The neuronal connections of the frontal ganglion of the cockroach Periplanetaamericana:a histological and iontophoretieal study[J].Cell Tissue Res,1978,193(2):353-371.
[13]MOORE D,LOHER W.Axonal projections within the brain-retrocerebral complex of the cricket,Teleogrylluscommodus[J].Cell Tissue Res,1988,252(3):501-514.
[14]RAND D,AYALI A.Neuroanatomyand neurophysiology of the locust hypocerebral ganglion[J].J Insect Physiol,2010,56(8):884-892.
[15]UDE J,AGRICOLA H. Synaptic connections of the nervus connectivus in the frontal ganglion ofPeriplanetaamericana L.(Insecta)[J].Cell Tissue Res,1979,204(1):155-159.
[16]SEHADOVA H,SHAO Q M,SEHNAL F,et al.Neurohormones as putative circadian clock output signals in thecentral nervous system of two cricket species[J].Cell Tissue Res,2007,328(1):239-255.
[17]常菊花,何月平。昆虫5-羟色胺受体的研究进展[J].安徽农业科学,2014,42(10):2913-2914.
[18]ANAVA S,RAND D,ZILBERSTEIN Y,et al.Innexin genes and gapjunction proteins in the locust frontal ganglion[J].Insect Biochem Mol Biol,2009,39(3):224-233.
[19]YE Hui,HUI Limei,KELLERBERGER K,et al.Mappingof neuropeptides in the crustacean stomatogastricnervous system byimagingmass spectrometry[J].J Am Soc Mass Spectrom,2013,24(1):134-147.
[20]AYALI A,ZILBERSTEIN Y.The locust frontal ganglion:a multi-tasked central pattern generator[J].Acta BiolHung,2004,55(1):129-136.
[21]RAND D,GUEIJMAN A,ZILBERSTEIN Y,et al.Interactions of suboesophageal ganglion and frontal ganglionmotor patterns in the locust[J].J Insect Physiol,2008,54(5):854-860.
[22]CAROL I M,BOOKER R.The role of the frontal ganglion in the feedingand eclosion behavior of the moth Manducasexta[J].J ExpBiol,1998,201(11):1785-1798.
[23]GILLOTT C.The role of the frontal ganglion in the control of protein metabolism in Locusta migratoria[J].Helgoland Mar Res,1964,9(1/2/3/4):141-149.
[24]VOLKER D,YVONNE K,ROLF K.The antennal motor system of the stick insect Carausius morosus:anatomyand antennal movement pattern duringwalking[J].J CompPhysiol A,2001,187(2):131-144.
[25]UCHIMURA K,Ai H,KUWASAWA K,et al.Excitatoryneural control of posterograde heartbeat bythe frontalganglion in the last instar larva of a lepidopteran,Bombyx mori[J].J CompPhysiol A,2006,192(2):175-185.
[26]MITSUMASU K,TANAKA Y,NIIMI T,et al.Novel gene encodingprecursor protein consistingof possibleseveral neuropeptides expressed in brain and frontal ganglion of the silkworm[J].Peptides,2009,30(7):1233-1240.