����ժҪ��Ŀ�ģ�SXT/R391Ԫ���Ǹ������Ծ����о����֡���֪��Ա���������ԽӺ�Ԫ�����壬���о�����ҪĿ����̽��SXT/R391Ԫ�����Ļ��������Ӧ�Խ������ɡ��������ռ����ݿ��м����ױ�����SXT/R391����Ԫ�����ݣ��������Ѳ����ϸ������������Ԥ��ü���Ԫ��������Ԫ�����Ļ���Ľ������ԡ������ͨ�������Ѽ���Ԥ�⣬�����83��SXT/R391Ԫ���Ļ��������С���52�����Ļ����У����㷢��22�������ڽ��������о��������飬����ֲ���SXT/R391Ԫ����4������ģ�顣���⣬��7�������м�����ѡ���źš����ۣ�������SXT/R391Ԫ�����Ļ���Ľ������ƣ�Ϊ�˽�ü���Ľ�������ɢ�춨�˻�����
�����ؼ��ʣ�SXT/R391Ԫ��; �������; ��ѡ�����;
����Bioinformatics Analysis on the SXT/R391 ICEs Core Genome
����ZHANG Qi ZHOU Jing ZHOU Jiang-Lin KONG Na HU Ming-Da PENG Xiao-Chuan LI Bei-Ping LIANG Long JIN Yuan REN Hong-Guang YUE Jun-Jie
����Beijing Institute of Biotechnology
����Abstract��Objective: To conduct adaptive evolution analysis of core genome of SXT/R391, which is one of the largest, diverse and well-studied family of integrating conjugative elements��ICEs�� among Gram-negative bacteria.Methods: We used all available SXT/R391 ICEs genomes to expound the genetic diversity at the genome level and then explore the evolutionary mechanisms contribute to its genetic diversity. Results: 83 complete genome sequences of SXT/R391 ICEs from different bacterial species and origins were collected. Among the 52 identified core genes, we found that 22 genes had recombination signals and 7 genes had significant evidence of positive selection. The results demonstrated that recombination contributed most to the diversity of SXT/R391 ICEs. Conclusion: We provide an overview of evolution in SXT/R391 ICEs, which will be helpful for understanding the evolutionary dynamics of the SXT/R391 family.
����ԭ������Ŀ��ƶ��Ŵ�Ԫ����mobile genetic element,MGEs����ͨ���Ӻϡ�ת����ת���ȷ�ʽ�鵼�Ŵ������ڲ�ͬ����ϸ����ĺ���ˮƽ������ת�ƣ�lateral/horizontal gene transfer,LGT/HGT����Burrus��2002���״ζ���Щ���Դ�Ⱦɫ���ϼ��в����Ͻ�����Ⱦɫ�壬ͨ���Ӻϵķ�ʽ��ϸ��ϸ��֮�����ת�Ƶ��Ŵ�Ԫ�������˹��࣬��֮Ϊ�����ԽӺ�Ԫ����integrating conjugative elements,ICEs)[1]��ICEs�����������ɾ������������Ӻ����������ƣ�����ͨ���Ӻϵķ�ʽ����ת�ƣ���������ͬ���ǣ����Dz����������ƣ��临�Ʒ�ʽ���º��ɾ������ƣ�ͨ�����Ͻ�����Ⱦɫ�壬������Ⱦɫ��һ����[2]��ICEs�������պʹ��������������Ӽ���ҩ�������ڵĶ������Ի������ϸ������ϸ���Ĺ��ܡ����dz���������ϸ���µ���״������ҩ�Ժ��²��Ե�[3]��
�����������ڶ���ϸ���з�����ICEs����Ŀǰ��û�й��ϵ�ICEs�������ŷ�f������˸�������ø�����������Լ����Ľṹͬ���Ե�������ICEs���з���ķ���[4]���������ַ������ѷ��ֵĴ�ICEs��Ϊ28�����塣SXT/R391��Ŀǰ��֪��ICEs���о����������ϸ��һ������[5]��
����SXTԪ�����緢����1992����ӡ�Ⱥ��ϼ������»��ҵ�O139Ⱥ���һ���MO10��Ⱦɫ���ϣ���Ԫ���ܹ������ϽӺϵķ�ʽ����������Ⱦɫ�塣SXTЯ��������ҩ�����ܹ��鵼���һ����Իǰ�����sulfamethoxazole,Su���������а���ण�trimethoprim,Tm������ù�أ�streptomycin,Sm������ù�أ�chloramphenicol,Cm������ҩ�ԡ���ʱ������Я���Ļ������Իǰ�����ͼ����а���वĿ��Զ�����ΪSXT[6]��R391��������70������ڱ�����Ϊ���迹����ù�ص��������������Ϊ�Dz�������Ⱥ����“IncJ”��ԭ��������20����ܶ��о�����R391������������“R����”���Ŵ�Ԫ��������SXT�߶����ƵĻ�����Ǽܽṹ�Լ��̶���Ⱦɫ�����/����λ�㣬������SXTʵ��������ͬһ���͵����ϽӺ�Ԫ������˽���һ��Ԫ������Ϊͬһ��SXT/R391���塣
����Ŀǰ���ֵ�SXT/R391Ԫ��������Ĵ�СΪ70��120 kb��SXT/R391Ԫ���Ļ�����������ı������Ϳɱ���������[5,7,8]��SXT/R391Ԫ���ı���������Ļ���鵼SXTԪ��������/�г����Ӻ�ת�ƺ�ICEs������������ĵ��أ�����һЩ���ػ���Ĺ���δ֪��SXT/R391�ɱ�������ı��������Ҫ���������Կ����ص���ҩ�ԡ��ؽ������ӿ��ԡ�����Ĥ�γɺ�ϸ���˶������ĵ��ڣ�����Ҳ���붾��-������ϵͳ������������ϵͳ������ø����������ø��[9]��
������Ϊ��ҩ��ҩ��multi-drug resistance,MDR����������壬SXTԪ���ھ���䲥ɢ������µ����ƾ������С�SXT/R391Ԫ���Ľ������������ɢ������ǿ��������������SXT/R391����Ԫ���Ľ������������ڷ�ֹSXTԪ����ɢ�������²��������о�����Ҫ��ʵ��ָ�����塣
����1 �����뷽��
����1.1 �����Ѽ�
����������“SXT/R391”��“ICE”Ϊ�ؼ�����PubMed��������ú��б���SXT/R391Ԫ�������ף�ͨ���Ķ�����ȡSXT/R391Ԫ����������Ϣ�����Ž���SXT/R391Ԫ����Ԥ�⣬Ϊ�˱�֤���������SXT/R391Ԫ�����У����Ǵӻ���������������Ϊ���������飨Complete Genome����Ⱦɫ�壨Chromosome��״̬��9632��ϸ������������������SXT/R391Ԫ����ͨ��BLASTp�������бȶԣ�ƥ�����а������������ģ�飨integration/excision module����DNA����ģ�飨DNA secretion module���͵���ģ�飨regulation module����3������ģ��ĵ��������ƶ�ΪDZ�ڵ�ICE�������һ�����������ڣ�70��130 kb�����ֱ���SXT/R391����Ԫ��������øInt�����ͷ���ϵͳ�Ӻϵ��͵��ص���SetR�Ļ�����Ԥ��û��������һ��������SXT/R391Ԫ�������գ�����ͨ������Ԥ����83��SXT/R391����Ԫ����
����1.2 �����
������ȡSXT/R391Ԫ����52�����Ļ������У���83��Ԫ����Blast����������ÿ��������83��Ԫ���ϵķֲ����������
����1.2.1 �������
����Ӧ�÷�������ѧ������RDP4[10]�е�RDP��GENECONV��BootScan��MaxChi��Chimaera��5�ֽ�Ϊ�����ķ������ֱ�̽��83��SXT/R391Ԫ����52�����Ļ������е������¼��������źš����У�GENECONV�IJ���g-scale����Ϊ1����������Ƭ���ڲ��Ĵ��䣻���������IJ�������ΪĬ��ֵ�������ġ�P≤0.01Ϊ����ͳ��ѧ�����Բ��졣
����1.2.2 ��ѡ�����
����ѡ�����߹���Datamonkey(http://datamonkey.org/��[11]����SXT/R391Ԫ�����Ļ������ѡ��ѹ������������ʹ��MUSCLE�������бȶԣ��ύ���м�ѡ������ģ�͡�����ѡ����4��ģ�ͣ������ЧӦ��Ȼ����REL�����̶�ЧӦ��Ȼ����FEL������һ���ȼ�������SLAC�������ЧӦ����ģ�ͷ�����MEME��������ωֵ[ω=dN/dS����ͬ���û���/ͬ���û��ʣ�]�ж�ѡ��ķ���ω>1�Ҳ�������ʱ�ж�Ϊ����ѡ��0<ω<1ʱΪ����ѡ��ω=1ʱΪ����ѡ��
����2 ���
����2.1 SXT/R391Ԫ�������������������ֲ�
����ͨ�������ռ���Ԥ����83��SXT/R391Ԫ���Ļ��������У���������������ȡ��54����ϸ��������������Ԥ���29����Ԫ����������������ķ���ʱ��Ϊ1967��2014�꣬�����ֲ�����21�����Һ͵�����Ԫ��������Խ25����������Ԥ���29��SXT/R391ICEԪ���У���12����������������δ��ȡ�ͱ���Ԫ�����������С�SXT/R391Ԫ���������㷺��ͼ1�������ֲ����ڽϴ�ƫ�ԣ��ڻ��һ����з��ֵ�������࣬������θ˾����������Ȥ���ǣ��ٴ����ֵ�SXT/R391Ԫ��Ҳ�������ڻ��һ�����������θ˾����ڻ����з��ֵ�SXT/R391Ԫ���������ˮ������SXT/R391�ľ��ֲַ������ȣ�˵��Ԫ�����������д��ںͱ��ؿ�����Ҫ����һ�����Ŵ�������
����2.2 ���Ļ������
����Wozniak�ȶ�13��SXT/R391Ԫ���Ļ���������˱ȽϷ���[7]�����ָ�Ԫ������52�����Ļ���������ȡ��SXT/R391Ԫ����52�����Ļ������У��Ի��ȫ��������Ϣ��83��SXT/R391Ԫ����ȫ���������н���ͬԴ����������Ԫ���ĺ��Ļ���ķֲ�����������ͼ2��
����83��SXT/R391Ԫ������һ������ij�ԱICEPmiChn3����Ԫ��ȱ��20�����Ļ�������13�����ӽӺϹ��ܡ�ȱ�ٵ�20�����Ļ���һ���µ���ת���Ӻ���ҩ������ɵĻ����ȡ����ʵ������������Ԫ���ĽӺϹ�����ȫɥʧ[12]�����ICEPmiChn3�Ѿ�����һ����������������ܵ�Ԫ������ˣ������ĺ��Ļ������������ų��˸�Ԫ����
�����������㣬52�����Ļ�������39����Ȼ������ȫ��Ԫ���С�������6���������1��Ԫ����ȱ�٣���Щ����ɳ�Ϊ�����Ļ���soft core)[13]������7������rumA��s024��s025��s026��traL/E/K����2����2������Ԫ����ȱʧ�����е�rumA��s024��s025��s026����SXT/R391Ԫ����ɢ�������С���ܻ��ϣ�ȱʧ��Щ������ܻ�Ӱ��Ԫ���Ĵ�����ɢ�����Ͻ��˵�������ױ�����13��Ԫ�����Ƶ�����52�����Ļ��������ϸ��صĺ��Ļ���������һЩ�������������Ļ���
��
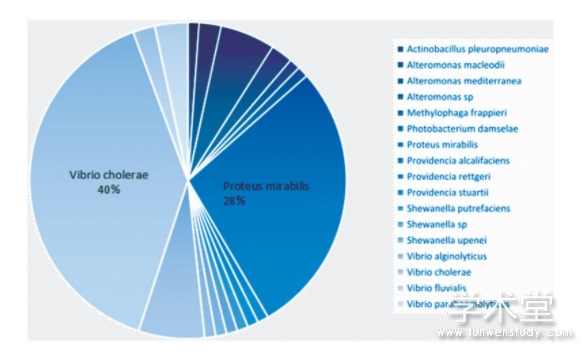
����ͼ1 SXT/R391�ľ����ֲ�
����ֵ��ע����ǣ�83��Ԫ��������ø������2�࣬�ֱ�Ϊint-1��int-2,2�������������Խ�Ϊ22%���ѽ�øxis����Ҳ��2�࣬�ֱ�Ϊxis-1��xis-2�����Ƿ�����Ԫ���У�int-1��xis-1Эͬ���֣�int-2��xis-2Эͬ���֡�
�������⣬��ͼ2���Կ�����traFHG��eex��setC��setD��s082��s083��s084��s086��setR�����ھ���Alteromonas macleodii str.MED6-ICE��SXT/R391Ԫ���г���������Ļ����ơ�
����2.3 ���Ļ�����ںϺ��ѽ�
�������˻����ƣ������������Ļ�������˻����ں�/�ѽ��źš�ȫ��rumB�������Ϊ1269 bp�����ײ���Ϊ423 aa�����û�����46��Ԫ������Ϊ2���������Ƭ�Σ�ͼ3����˵��rumB�������˻����ں�/�ѽ⣬������λ�㸽��������Ƶ���IJ����йء�
�������⣬trh F��tra W��һЩԪ����Ϊ2����������Ԫ��Proteus mirabilis MD20140901(KX243408���У�trh F����[54557..55069]������513 bp�����ײ���171 aa;tra W����[55080..56204]������1125 bp�����ײ���375 aa�����磬��Ԫ��Vibrio cholerae O37MZ03(JQ345361���У�trhF����[26846..27358]������513 bp�����ײ���171 aa;traW����[27369..28493]������1125 bp�����ײ���375 aa������Ԫ��Providencia stuartii ATCC33672(CP008920���У���Ӧ��2�������ں�Ϊ1�������ںϻ������[1904626..1906272]��������λ�ã�������1647 bp���ںϵ��ײ��549 aa��Ϊtype-F conjugative transfer system pilin assembly family protein(AIN64681.1����
����

����ͼ2 ���Ļ���ֲ����
��

����ͼ3 rumB�����ڲ�ͬԪ���еķֲ�
����2.4 ���Ļ�����������
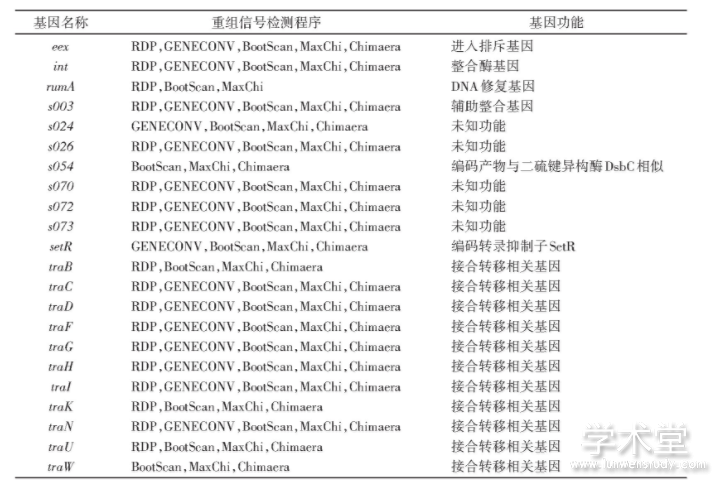
����Ԫ��֮��������ܵ����γ��µ��Ӻ�Ԫ���������¼���SXT/R391Ԫ�������Բ����Ķ���֮һ�����о�������SXT/R391Ԫ�����������ڻ�������顣������˷����ڻ���������ˮƽ�������ܷ������ǻ���ˮƽ��sub-gene level����Ϊ��̽��SXT/R391Ԫ�����ǻ���ˮƽ�����¼������Ǽ����SXT/R391Ԫ��52�����Ļ���Ļ��������顣��52�����Ļ����У���22�����Ļ�����3�ּ�3�����ϵķ����п��Լ����Ե������źţ���1����
����ͼ4�ú�ɫ��������������SXT/R391Ԫ���������ϵ�λ�ã���Щ�����漰Ԫ�����Ļ����ȫ��4�����蹦��ģ�飬�����������ģ�顢DNA���Ƽ��ӹ�ģ�飨replication/DNA processing module����DNA����ģ��͵���ģ�顣����ģ�飬�����ͷ���ϵͳ��type��secretion system,T4SS���йػ���صĴֻ����������źš����⣬����һЩ���������źŵĺ��Ļ���Ϊδ֪���ܻ���
����2.5 ��ѡ�����
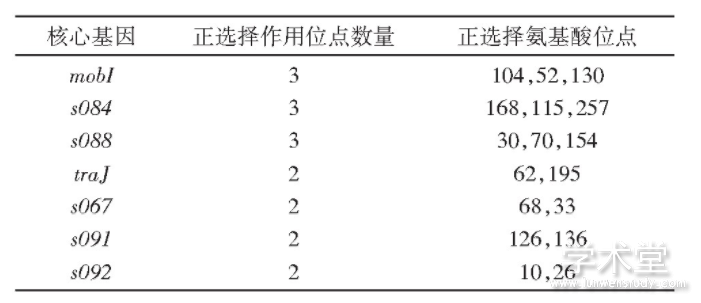
������ѡ��positive selection��Ҳ��������������ж������Ŵ����������������������Ӧ�Խ����з��ӹؼ����á����Ǽ����SXT/R391Ԫ���������л������ѡ���źš�Ϊ�˱��������¼�����ѡ�������ɸ��ţ�����ֻ��δ���������¼��ĺ��Ļ����ϼ����ѡ��������ʾ����ȥ���Ϸ�����22����������ĺ��Ļ���SXT/R391Ԫ����û�������źŵ�30�����Ļ�������7������ѡ���źţ���2������7�������е�mobI��traJ����SXT/R391Ԫ���Ӻ�ת�����走�ף�����5���������δ֪���ܵ��ף��Ҳ���SXT/R391 ICE��С���ܻ��ϣ�minimal functional SXT/R391 ICE gene set���У����Ƕ�SXT/R391Ԫ�������ԵĹ����д���һ���о���
������1 ���Ļ��������źż��
��

����ͼ4 22����������ĺ��Ļ�����SXT/R391Ԫ���Ǽ��ϵ�λ��
������2 ���Ļ������ѡ����������λ��
��������I-TASSER������mobI��TraJ�������ĵ�������ά�ṹԤ��[14]����Ԥ�����ά�ṹͼ��ͼ5���п��Կ�����mobI�������ĵ�������ѡ��λ��52��104�л�����α�����ṹ�Ķ�ͷ��130�л�λ����һα�����ṹ�ϣ�TraJ�������ĵ�������ѡ���λ��62�л�������α�����ṹ�Ķ�ͷλ�ã�195�л�λ��α�����ṹ�ϣ�����Ԥ��õ���191��210�л�Ϊһ����Ĥ�ṹ����ѡ��λ��195�л����ڸýṹ���ڡ�����λ�������ĵ��ṹ�����ת�������йأ���Щλ���ܵ�ѡ��ѹ��������Ԫ����ת����ɢ��ء�
��
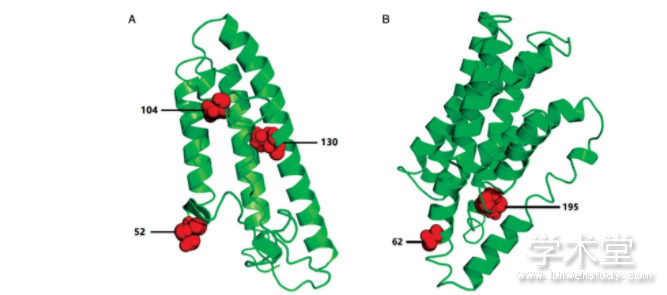
����ͼ5 mobI����A����traJ����B�����뵰����ά�ṹ����ѡ��λ��
����3 ����
������ΪMDR��������壬SXTԪ���ھ���䲥ɢ������µ����ƾ������С�SXT/R391Ԫ���Ľ������������ɢ������ǿ��������������SXT/R391����ICEs�Ľ���������Ǩ��ģʽ����ɢ���ķ��ӻ��ƣ������ڷ�ֹSXTԪ����ɢ�����²��������о�����Ҫ��ʵ��ָ�����塣SXT/R391Ԫ���ĺ��Ļ��������Ԫ���ĸ��ƺ�ת�ơ��������ѡ�����������������2�ֶ������Ŵ����������������������Ӧ�Խ����з��ӹؼ����á����о��ռ���Ŀǰ���ֵ�SXT/R391Ԫ�����Ժ��Ļ���ķֲ��Լ����Ļ�����������ѡ������������������˷�����
�������о�������ͬԴ�������ѡ����SXT/R391Ԫ�����Ļ���Ľ��������з�������Ҫ���á����Ƿ�����22�����Ļ����ڽ����о�����ͬԴ���飬������Ԫ���൱һ���ֺ��Ļ�����Ŵ����졣���⣬��7�����Ļ��������ѡ���źţ�����2������mobI��traJ��Ԫ���ĽӺ�ת���йء�Ԫ�������������ѡ����춼��Ϊ��SXT/R391Ԫ���ʺ϶ȵ��������Ӷ�����Ӧ��̬�����ı仯�����ϣ����Ƿ�����SXT/R391Ԫ�����Ļ������������Ӧ�Խ�����Ϊ�˽�SXT/R391Ԫ������ɢ���������ɵ춨�˻�����
���������
����[1] Burrus V, Pavlovic G, Decaris B, et al. Conjugative transposons:the tip of the iceberg[J]. Mol Microbiol,2002,46(3):601-610.
����[2] Johnson C M, Grossman A D. Integrative and conjugative elements(ICEs):What they do and how they work[J]. Annu Rev Genet, 2015,49(1):577-601.
����[3] Rodríguezblanco A, Lemos M L, Osorio C R. Integrating conjugative elements as vectors of antibiotic, mercury,and quaternary ammonium compound resistance in marine aquaculture environments[J]. Antimicrob Agents Chemother, 2012,56(5):2619-2626.
����[4] Bi D X, Xu Z, Harrison E M, et al. ICEberg:a webbased resource for integrative and conjugative elements found in bacteria[J]. Nucleic Acids Res, 2012,40(D1):D621-D626.
����[5] Carraro N, Burrus V. Biology of three ICE families:SXT/R391, ICEBs1, and ICESt1/ICESt3[J]. Microbiol Spectrum, 2014,2(6):MDNA3-0008.
����[6]����,������,����Ⱥ.ϸ�������ԽӺ�Ԫ��SXT/R391�о���չ[J].����ѧ��, 2014,54(5):471.
����[7] Wozniak R A F, Fouts D E, Spagnoletti M, et al.Comparative ICE genomics:insights into the evolution of the SXT/R391 family of ICEs[J]. PLoS Genet, 2009,5(12):e1000786.
����[8] Zakharova I B, Viktorov D V. Integrative conjugative elements(ICEs)of microorganisms[J]. Mol Genet Microbiol Virol, 2015,30(3):114-123.
����[9] Garriss G, Burrus V. Integrating conjugative elements of the SXT/R391 family[M]//Roberts A P, Mullany P.Bacterial integrative mobile genetic elements. Landes Bioscience, 2013:217-234.
����[10] Martin D P, Murrell B, Golden M, et al. RDP4:detection and analysis of recombination patterns in virus genomes[J]. Virus Evol, 2015,1(1):vev003.
����[11] Pond S L K, Frost S D W. Datamonkey:rapid detection of selective pressure on individual sites of codon alignments[J]. Bioinformatics, 2005,21(10):2531-2533.
����[12] Bie L, Wu H, Wang X H, et al. Identification and characterization of new members of the SXT/R391 family of integrative and conjugative elements(ICEs)in Proteus mirabilis[J]. Int J Antimicrob Agents, 2017,50:242-246.
����[13] Bioteau A, Durand R, Burrus V. Redefinition and unification of the SXT/R391 family of integrative and conjugative elements[J]. Appl Environ Microbiol, 2018,84:e00485-18.
����[14] Yang J, Zhang Y. I-TASSER server:new development for protein structure and function predictions[J]. Nucleic Acids Res,2015,43(W1):W174-181.