
������Ѫ�쵰����������ϵͳ�о�����������ѧ�������Ҫ�ɹ�֮һ����ṹ�빦�ܵĹ�ϵҲһֱ����ؽ�ѧ��ע���ص㡣�ڽ�һ�������������о��У����Ƕ���Ѫ�쵰�ķ��ӽṹ����������������ѧ���ܶ����˽�Ϊȫ�����ʶ; ���Ƕ������������Ѫ�����ڽ��������ǰ�����ؽṹ�仯��Ŀǰ��Ȼ���ڶ������飬���а���: Ѫ�������ĵ����������ں������ӽ�ϵĹ������Ƿ����˵���ת��?
������������Ѫ�������Ƕ���λ���Dz���λ? ������״̬���Ƿ���ˮ����ռ�ݵ� 6 ����λ��? ���Ľ���о���չ����Щ��������ܽ���������ڼ����Ѫ�쵰�ṹ���ܵ����⣬������ѧ���ݣ�����Ѫ���������������ƺϳ��ṩ�ο���
����1 Ѫ���صĽṹ�빦��
����Ѫ�쵰���ڸߵȶ����������������������á�Ѫ�쵰�ף�1�ݾ��е��ļ��ṹΪ4 ���ǻ����ɵ���Դ�ľ��� α2β2������ÿ�������ǻ������һ��Ѫ���ظ������ҽ�������ڵ��ײ������ɵ���ˮ����( ͼ1) ��
����
��������״̬��Ѫ���ط�����߲��������������λ�γɡ���Ȼ��ͬѪ���ط��ӵ�߲�����������ṹ��ͬ�����Է�Ϊ�����칹�壬�������û��� 4 ����ԭ�ӽ�����λ�ķ�ʽ��һ�µġ�ͬʱ�������� 87 λ( α�ǻ�) �� 92 λ( β �ǻ�) ���鰱��л��ϵ�����ԭ�Ӵ�߲����ƽ���Ϸ�������������λ�����ɵ� 5 ��λ��( ͼ 2) ��
����
����ʵ��֤����������Ѫ�����У��������Ӱ뾶�ϴ�( 92pm) ������Ƕ��߲�����ڣ�����߳�߲����ƽ��60pm������������λ�����������Ӱ뾶��С�� 75pm ������߲����ƽ�档�ڽ�������ӹ����л����������������ӵĴžر仯��������״̬�£�Ѫ����������˳����; ��������״̬�³ʷ����ԣ�1�ݡ�
��������Ѫ��������ǰ�������Ӱ뾶�ʹ��Ա仯��ԭ����ڶ������飬����Ѫ���������������������Ϲ������Ƿ����˵���ת��������Ҫ��һ������˹������( M?ssbauer spectroscopy) ֻ��ʾ������������������֮����ǿ�Ĺ������ã��������Զ������ӵ�����̬���������˵����2�ݡ����⣬I�� ���й۲쵽��O—O���ĺ���������Ƶ��������������϶����ڷ����3�ݣ�������ȷ��һ����O—O���ļ���Ϊ1�� 5���볬�������ӵļ�����ͬ��4�ݡ���aman ����Ҳ�ṩ��һЩ���ݣ�5�ݡ�����������������������������λǰ���ԭ�ӹ�������ӹ���������飬���������Լ����������������ǵ�������Ȼ�ᷢ���仯����û��ֱ��֤�ݵ�����£�ֱ�ӶԱ���λǰ���������ʺ���ѧ���ݣ��Ӷ��ó�֧��ij�ּ���Ľ����Dz�����ֿɿ��ģ�6�ݡ�
����������������飬һ�������������ϵ������ӵ���λ��ʽ�����Ƕ��仹�Dz���; ��������Ѫ���������������� 5 ��λ��������ˮ������λ�γ� 6 ��λ����������˲�ͬ����������ģ�ͣ���ͼ���������۽��н��͡�
����2 ����ģ���о�
������������ʵ��֤�ݱ���������Ѫ�쵰��ת��Ϊ����Ѫ�쵰�����д���ijЩ���ӵĽṹ�仯�͵������Ź��̣����Ƕ�Ѫ�������������Ϲ��̺ͽṹ����˶�������ģ�͡�
�������磬Pauling ���þ���ۼ����۽���ģ�ͣ�7-9�ݶԴ˹��̽���˵��: O2�Զ��䷽ʽ���������ӽ�ϣ�����һ����ԭ�ӵ�һ�Թ¶Ե���������λ��δ��������ת�ƣ����������� 5 ��λ������״̬ת��Ϊ 6 ��λ������������뾶��С�����ڵ�����ԣ�˳����ת��Ϊ�����ԡ�Ѫ������һ����̼��ϵIJ���������Ƶķ��ӽṹ������˵��Ѫ�������Ϻ����������һ�ֹ����壬�������-��֮��ļ��в���˫������( ͼ 3a) ��
����������Weiss �������ת��ģ�ͣ�10-11�ݣ���Ϊ��Ѫ������˫����Ϻ����ĵ��������� Fe( ��) ����ת��һ�����Ӹ� O2�γɳ��������� O��2��������ɰ뾶��С�ĸ������� Fe( ��) ��Ȼ���������������������λ����Ȼ�������ĸ���������һ��δ�ɶԵ��ӣ�������˳���ԣ�������ͬ������һ��δ�ɶԵ��ӵij�����������λ��Ϻ�����ԣ����Ե�˳������ʧ��������ʾ������( ͼ 3b) ��
��������֮�⣬Griffith ���������һ����������ģ�ͣ�12�ݡ���ͨ��������ԭ�����������ӵĹ������ó�������λģʽ�Ȳ��Ǵ�ֱ���乲���壬Ҳ������б���乲���塣�������״̬����������Ϊ O2��ƽ����߲��-����λƽ�������λ�ģ����������ϣ������������ӿյ� d2sp3�ӻ������ O2�� π2p����ص��γ� σ �������������ӵ� 3dxzԭ�ӹ���ϵĵ��ӿ��Ժ� O2�� π*2p�չ���γɷ��� π ��( ͼ 3c) ��
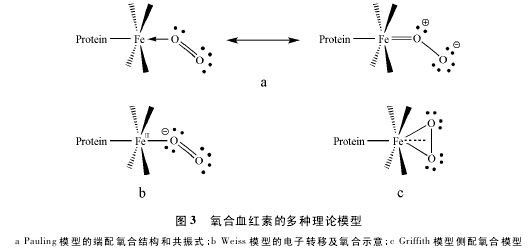
���ϣ����ڴ��� 3 �ֲ�ͬ����ģ�ͣ�����Լ�������ֱ����������
����3 ����һ: ����Ѫ�����������ӵ�����̬����
�������ȣ����ڽ��˫��ǰ��Ѫ��������������������̬�ı仯������ڴ������顣
�����Թ��ϵĴ�����뾶�仯����ʵ��Pauling ���۽���Ϊ Fe( ��) ���ӵĵ�������״̬�ĸ�[]���ϣ����ڴ��� 3 �ֲ�ͬ����ģ�ͣ�����Լ�������ֱ����������3 ����һ: ����Ѫ�����������ӵ�����̬����.���ȣ����ڽ��˫��ǰ��Ѫ��������������������̬�ı仯������ڴ������顣�Թ��ϵĴ�����뾶�仯����ʵ��Pauling ���۽���Ϊ Fe( ��) ���ӵĵ�������״̬�ĸı䣬������������ǵ�����Ҳ����ϣ�13��; Weiss ģ������Ϊ��( ��) ������Ϊ��( ��) �����������ǵ�������״̬�ı仯������ԭ������̬�����߱�Ȼ�����Ű뾶�ļ�С�ʹ��Եĸı䡣ǰ���ᵽ�� I�� ���й۲쵽O—O���ĺ���������Ƶ�ʶ�Ӧ��O—O���ļ���Ϊ 1�� 5���볬�������ӵļ�����ͬ��14�ݡ����⣬X ���߹��������( X-ray photoelectron spectroscopy) �о�������������ԭ�Ӿ���һ��ԼΪ 3�� 2 ������̬��15�ݡ�Durham���ˣ�6������ X �������ս��߽ṹ����( X-ray absorption near edge structures) �о����������������Ϲ���ǰ����ܼ��仯�����֤�������������Ϻ���Ч����������ˡ���Щ���ܹ��� Weiss ģ�ͺ����ؽ��͡�
�������� Pauling ģ�ʹ���һЩ������ͨ������������������ת�俴���ܹ����ʹ��Եı仯��������ͬʱҪ��������������������������ƽ�е�����̬ת��Ϊ������Եĵ���̬������һ�������ϲ����Ĺ��̡�ͬʱ���� Pauling �����У���Ϊ����λ֮��ʹ�����峡�ľ��峡���������ߣ��Ӷ�������������������˳���Եĸ�����( t42ge2g) ת��Ϊ�����Եĵ�����( t62ge0g) ������ʹ��뾶��С����������������У���λ�������ܵı仯���Լ��ߵ�������ת��뾶��Ӱ�죬��ȱ��ֱ������֤�ݵġ����֮�£�Weissģ������������ת��Ϊ�������ӣ����Ӱ뾶��Ȼ��С�Ľ�����ֱ��������
�������ǣ�Ŀǰ�� Weiss ģ�����ķ����ǵ��͵ĸ���Ѫ����ȴ���߱������������ܡ�������һ�㣬������Ϊ�����������ͣ�һ����Ϊ�� Weiss ģ���У�����������λ��ʵ�����dz��������ӣ������ǿ�����ֱ�Ӵ��ڵ�˫�����ӡ��ʶ�����һ�㲻����Ϊ�� Weiss ģ�͵�֤�ݡ����⣬ʵ���16�ݱ�������Ѫ�쵰���з���� �� Ѫ �� �� �� ¶ �� �� �� �� ʱ���� �� �� �� �� �� �� Ѫ �� �� �� �� �� �� �� λ���� �� �� �� ��Fe( ��) -O2-Fe( ��) ���Ѫ���������������������ΪѪ���ظ�����ͬʱ���鵰�����γɵ���ˮ�ṹ���Ա���Ѫ���������ڿ������������в������������ѱ����ϡ�������ǿ�����Ϊ���� Weiss ģ���У���Ѫ�����������ӵ������ӵ�һ������ת��ֻ������������λ���У������ǿ���ġ������������У��ܵ���Χ���ṹ�ı�����������λ���ѵ������ӣ�Ѫ���ػָ�Ϊ�������ӡ��ɼ���Weiss ģ�Ϳ��Խ�Ϊ�����ؽ������д������û�����Բ�����֮����
��������Ѫ�����������ӵ�����̬֮���ڽ�ѧ��ϵ���Ƿdz������ģ���Ϊ�漰Ѫ�쵰�Ŀ������������ϻ����Ƿdz���Ҫ�ġ���ʷ�� Pauling ģ�����Ź㷺��Ӱ�죬Ŀǰ������ܶྭ��̿����ж���ͬ��һ�۵㡣���磬Miessler��17���ڡ�Inorganic Chemistry���в���Ѫ�쵰�Ĵ��ڿ��Ա���Ѫ������ʹ����������Ϻ��Ա��� Fe( ��) ��ʽ������������Ϊ Fe( ��) ����������18���ڡ��ߵ����ṹ��ѧ����Ҳǿ������Ѫ�쵰���У�O2����� Fe( ��) ����������Ϊ Fe( ��) ������һЩ�µ���ѧʵ����о���Weiss ģ��Ҳ�õ�Խ��Խ���֧�֡�Cotton��19���ڡ�Advancd Inorganic Chemistry����Ҳ�ᵽ��������������Ѫ��������Fe( ��) ������; ���Ϻ��Ϊ Fe( ��) ��������ͨ���볬�������ӽ�϶�������Գ��ַ����ԡ����ӽ������䡶��������ѧ���̲ģ�15����֧�����Ϲ����е� Weiss ģ�͡�
����4 �����: ˫����λ��ʽ
�������� O2���������ӵ���λ�������Ͷ���( bent end-on) ���Dz���( side-on) Ҳһֱ�������顣��Griffith ģ���У�O2��Ѫ����������ԭ����λ��ʽΪ����; ���� Pauling ģ�ͣ�7-9�ݺ� Weiss ģ�ͣ�10-11���У�O2�������Ͷ��䷽ʽ������ϡ�ģ�ͻ������Ѫ�쵰�ס����쵰�� X ���߾����������ݶ�֤ʵ˫�����������Զ��䷽ʽ�������һ����б�ǡ��� M?ssbauer ��Ҳ֤�� O2�Ȳ���ƽ��Ҳ���Ǵ�ֱ��Ѫ����ƽ�档
������������Ի��ȷ�ĵ�����ṹ���ݣ�һϵ����Ѫ�쵰�����弸�ι������Ƶ�ģ�ͻ����ﱻ���������о������ʽṹ�빦��֮��Ĺ�ϵ��20�ݡ�Ѫ�쵰�������������������Ͷ���ṹ�����ڼ����������( “picket fence”complex) ( ͼ 4) �б��۲��21-22�ݣ��������������ϼ��쵰�ף�23�ݼ�����Ѫ�쵰�ף�24�ݵľ���ṹ��Ҳ�۲쵽��������λ��ʽ( ͼ 5) ������Ѫ�����е�Fe—O—O���ڲ�ͬ���ֵ�ͬԴ�����ʼ�ģ�ͻ����������ض��Ƕȣ�Fe—O—O���ļ����� Fe( TpivPP) ( 2-Melm) �������������Ϊ 129°; ������������Ѫ�쵰����Ϊ 170°������Ϊֱ���ͣ�25��; �ڼ��������ϼ��쵰����Ϊ 115°��26��; �ڼ���������Ѫ�쵰����Ϊ 156°�����ڼ�����������������Ѫ�쵰��֮�䡣
��������ǻ�Ĵ�С��˫���뵰�����ض�������֮��������γɻ�Ӱ��Fe—O—O���ǡ����磬���ݿռ�λ���жϣ�����Ѫ�쵰�����ϼ��쵰���� α-�ǻ� E7 λ���鰱���ϵ� Nε�� O �ļ���� 2�� 7?���������γ� Nε? H ��O ������������ǿ����α-�ǻ� E7 λ�鰱����˫�������λ�ý�Ӱ��Fe—O—O���ǡ�
����5 ������: �������������
����������Ѫ�쵰���У�Ѫ�������γ�6 ��λ�ṹ������ CO��NO ��С���Ӽ���ȡ�� O2��Ѫ�������γ����Ͷ���ṹ��27�ݡ��������������Ѫ�쵰���У�ˮ�Ƿ���Ѫ��������λҲС�����顣�ֽ̲���������Ѫ�쵰������������Ϊ 5 ��λ����ΪѪ������������ˮ���ṹӦ������ˮ����λ���; ��Ҳ���о���28�ݱ���������Ѫ�쵰���У�ˮ���ӿ�����ΪѪ���������������������λ���������������Ѫ������Ϊ 6 ��λ�����Ѫ������������ǰ���� 6 ��λ����ֻ��ˮ���ӱ�������ȡ��������ѽ���Ѫ�����������Ϲ����еľ�ṹ�仯������ˮ������Ѫ����������λ�����Ŀǰ���õ�Ѫ�쵰��Ѫ����ģ�ͻ�����ľ���ṹ��ʾ������ˮ�������Ľ�������λ��19�ݡ��� Priedman ���ˣ�29�����о� Scapharca �Ķ���Ѫ�쵰�ṹʱ���֣��ڴ�Ѫ�쵰���ǻ���������ˮ�����Ŵؽṹ�����ݶ���Ѫ�쵰����ṹ����������Ѫ�쵰��Ѫ����������ˮ������λ�������ö���ͻ�䡢�ı���ѹ�ķ�����һ���о�ˮ������λ��Ѫ�쵰���ܵ�Ӱ�죬���ֽ� Scapharca Ѫ�쵰�� 72 λ�հ���ͻ��Ϊ�Ӱ����������Ѫ�쵰����������� 40 ��; �����һ���������Ҫ�������: ͻ���Ѫ�쵰���е�Ѫ��������ˮ���ӵĽ�����������ˡ����⣬������ѹ��������Ѫ�쵰�����������ԣ���˵��ˮ���Ӻ���������Ѫ��������λʱ���ھ�����ϵ����Ѫ�쵰����Ѫ������������ˮ���Ӵ������Ƶ�����λ����

��������������Ŀǰֻ�ڼ�����Ѫ�쵰�����й۲쵽��ˮ��Ѫ����������λ��һ����Ϊ������ʱѪ������Ϊ 5 ��λ����Ѫ����������ˮ������λ��
����6 ����
��������Ѫ�쵰����Ѫ���ص����Ϲ��̼�����ǰ��Ľṹ�仯�����Dz����˶��������ͻ�ѧ�ֶν����о�������˷ḻ�����ݡ����ڴˣ���������� Pauling ģ�͡�Weiss ģ�͵Ȳ�ͬ������ģ����ͼ����Ѫ�쵰�����ϻ��������Ƕ������ṹijЩ�������ʶ��Ȼ����һЩ���顣����ͨ�������Ƚϼ�������ģ�ͣ��������о������½�չ����Ŀǰ��ѧ�������������������������ܽᣬ�����ؿ������������Ϲ����������ӵ�����̬���⣬������Ϊ Weiss ������ Fe( ��) �볬�������ӵ���λģ�ͱȽϺ���; ���� O2��λ��ʽ�����飬Ŀǰ�ѻ������������ṹ������Ϊ�����͵Ķ���; ��������Ѫ�����������ӵ���λ����ʵ���鲻����Ϊ�� 5 ��λ����ijЩ�о���������������£�һЩ���ֵ�Ѫ����������ˮ���鰱���߲�����γ� 6 ��λ�ṹ��ֵ�ù�ע��
������ �� �� ��
������1�� ��˹�ң�Ҧ���죮 �м�����ѧ�� ����: ������ѧ�����磬2003
������2�� Lang G��Marshallproc W�� Proc Phys Soc��1966��87: 3
������3�� Momenteau M����eed C A�� Chem ��ev��1994��94: 659
������4�� Barlow C H��Maxwell J C��Wallace W J��et al�� Biochem Biophys ��es Commun��1973��55: 91
������5�� Tsubaki M��Nagai K��Kitagawa T�� Biochemistry��1980��19: 379





