
�����ռ��������Dz�����ֵ���Ҫԭ��֮һ,�ռ��ֶ���Դ�����������ķֲ���ʽ����Ͼ��о���������,����̬ϵͳ��һ���dz���Ҫ�Ľṹ���ԡ��ռ��������о�����ʶ��ͬ�߶��ϵ���̬ϵͳ�ṹ�����ܺ��̾�����Ҫ���������塣���,̽�ֲ�ͬ�߶��Ͽռ������Եķ�չ��ЧӦ,�о��ռ������Զ����P��������̵�Ӱ������,��ȷ�������Ϳ��ƿռ��ֵľ������Ӻͻ���,������ֵ���̬ѧЧӦ��������̬ѧ���̵������,��Ϊ��̬ѧ�о��е��ص����⡣
�����ռ������ԵĶ����������������÷ֲ���ָ�������ֲ���Ϸ����������Ͷ����Բⶨ���϶ȷ����������۸�ֶ����Բⶨ�ȴ�ͳ����,��Щ�����ڹ������о��еõ��˹㷺��Ӧ��; ��ͳ��ѧ�ͷ��η����ȷ���������С�߶ȿռ�ṹ���о���Ҳ�ܵ���������Ӣ������ѧ�� Taylor1960 �����о�������Ⱥ�ֲ��з��ֵ��ݳ˷�����,���ձ�ѧ�� Shiyomi��Ӧ���ڲݵ�ֲ��Ⱥ��ռ������Ե��о���,�õ��˺ܺõ�Ч����Ŀǰ,����ѧ�߶�Ⱥ��ռ������Ե��о��Լ����ڴ�ͳ�ķ�����������,��ͳ���鷨���������ɼ������ֵ��鼰�ռ������Եķ�����,���ں�ʱ�����Ѷȴ��ȱ��,���ݺ�������ij���ǡǡ�ֲ��˴�ͳ���鷽���IJ��㡣���,���ַ���ֵ���ڹ����о��е��Թ㷺�ƹ�Ϳ�չ��
�������ױ�����ԭ���й�������Ҫ������ҵ�������غ���̬��ȫ����,�����������ڲ������Ŀ�����������,��֮����仯��Ӱ��,��ԭ�˻�����Ӿ硣������,����ѧ�ߴ�����ɳĮ���������Ĺ�ϵ��ֲ��Ⱥ��ռ��֡���������̬����������Ա仯�ȷ��濪չ����Ӧ�о�,����Բ�ԭȺ�����ö�������ѧ����,����̽�ַ���Ӱ����С�߶���ֲ��Ⱥ��������ɽṹ���ʻ��ֲ�������о��в������
�������������ɹź��ױ�����ԭ��Ϊ�о�����,�����ݺ���������Բ�ͬ�����ݶ���Ⱥ��ռ������Էֲ����������ֶ����Ե�Ⱥ��ѧ������չ�����о�,ּ��Ӧ������ͳ��ѧ��������,�Բ�ԭȺ�������Էֲ��������ж�������,�����о���Ⱥ��Ⱥ��ռ�ֲ�����,����Ϊָ�����������ͺ������ò�ԭ��Դ�ṩ��Ӧ���������ݡ�
����1�� �о����ſ�
�����о�����Ϊ���ɹź��ױ�����ԭ��³������Ŀ�����é(Stipa krylovii) Ⱥ��,���ص���λ��47°55'03. 3″ ~ 49°14'43. 9″N��116°40'12. 7″ ~ 117°23'24. 1″E�������´���ɺ���������,�����ˮ��Ϊ200 ~ 300 mm,��ƽ������ - 5 �� ~ 2 �� ,��˪�� 85~ 155 d,≥10 �� ������� 2 320 �� ,���� 125 d ���ҡ�Ⱥ��������Ϊ������é(Stipa krylovii) �����(Leymus chinensis) �������Ӳ�(Cleistogenes squarro-sa) ��,�ش�������������ҪΪ��������
����2�� �о����������ݴ���
����2. 1 Ұ�����
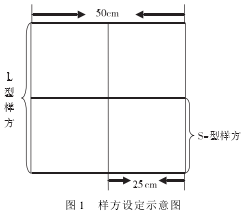
�������� GPS ���ж�λ,������������������ 3 �������ݶ���,�ֱ�ѡ����͵ص�,������һ�� 50m ������; ���������� 50 cm × 50 cm ��������(��� L ������,����100 ��) ���е���; ÿ�� L �������ֻ���Ϊ 4 �� 25 cm ×25 cm ��С����(���S ������,���� 400 ��) ,���� S �������ڵ��������(��ͼ 1) ��
����
����2. 2 ���ݴ���
����2. 2. 1 ��Ⱥ�ռ������ԵIJⶨ
������������ͳ��ѧ��β - ����ʽ�ֲ�(bata - bino-mial distribution) ���ݴη�(power - law) ���вⶨ�������Ⱥ�ռ�ֲ���֡���ν�ݺ�������,����x = log(p(1 - p) / n) ��Ϊ x ��,y = log(v / n2) ��Ϊ y��(������ȡ�������� 10 Ϊ��) ,�����ֵ����ж�Ӧ�� x��y ��ɢ��ͼ,y ������ x �Ļع鷽��ʽ��ʾ�ľ��鷨��,����: n Ϊһ�� L - ����������ֵ� S- ��������,n = 4; v Ϊʵ�ʹ۲���ij���ֳ������ķ���; p(1 - p) /n Ϊij����Ϊ����ֲ�ʱ,�������ķ���; piΪ���� i �� S �������г��ֵ�Ƶ�ȡ�
����Ⱥ���и����ֵij���Ƶ����ռ�ֲ�������ָ���˻���ƽ��ֵ,����“Ⱥ��Ŀռ�ֲ�������ָ��”��Ϊ δc,�� δc ����Ⱥ��IJ���һ�Ի������Գ̶�,���� δc = ∑(pi× δi) /∑pi���㡣
������һ�������� ε ��ʾ����ƫ���ݺ�������ֱ�ߵij̶�: εi= yi- y,�� εi���������� i ��ʵ�ʷֲ��ķ���Ķ���ֵ yi������ֲ�ʱ xi�Ļع鷽��yi= a + bxi֮��IJ�ֵ,��ʾ�� i �Ŀռ���Բ���һ�Եij̶ȡ�ε =0,������ i �Ŀռ�ֲ���Ⱥ��������Ƚ�,����һ�µ�������(����һ��) ; ε >0,������ i ��Ⱥ��������Ƚ�,���иߵ�������; ε <0,������ i ��Ⱥ��������Ƚ�,���е͵������ԡ�
����2. 2. 2 ���ֶ����Լ���pi= ni/ N,ʽ��,niΪ�� i ���ֵĶ��ָ��; N Ϊ�����и������ֶ��ָ����ܺ͡�
����Simpson ��̬���ƶȶ�����ָ��:D = ∑ (pi)2��
����Shannon - Wiener ������ָ��:H = - ∑pilgpi��
����2. 2. 3 ���־��ȶȵļ���E. Pielou ���ȶ�ָ��:E = H / ln(S) ,ʽ��,H Ϊ Shannon - Wiener ������ָ��,S ΪȺ����������Ⱥ��Ŀռ�������ָ��: δc =Σ(pi× δi) /Σpi��
����3�� ��������
����3. 1 ��ͬ�����ݶ���Ⱥ��������ɵı仯
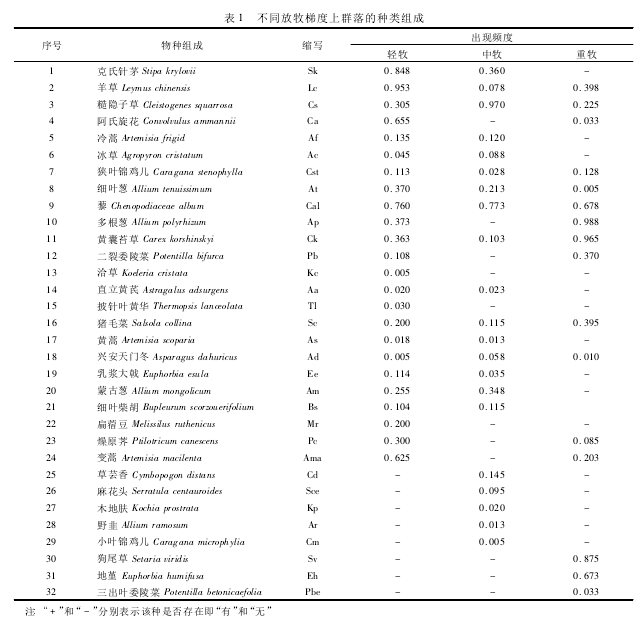
������ͬ�����ݶ���Ⱥ���������ɼ���1���ɱ�1 ��֪,����Ⱥ�乹�����ֵij���Ƶ��ֵȷ�����������,��ȷ����ݶ��ϵ�Ⱥ������ݺͿ�����é�ȹ�24 ���������,������ݺͿ�����é����Ƶ�����; �жȷ����ݶ��ϵ�Ⱥ���ɲ����Ӳݡ�������é���ȹ� 21 ���������,�����Բ����Ӳݳ���Ƶ�����,���ֳ���ԽϺõ�������; �ضȷ����ݶ���,Ⱥ�������ɶ���С�����̦�ݺ��ȹ�16 ���������,����кͻ���̦�ݱ��ֳ��Ϻõ������ԡ��ڲ�ͬ���÷�ʽ��,Ⱥ��ṹ���Ըı�,���Ⱥ������������仯����Ⱥ���е����Ƶ�λҲ�������ʽ�IJ�ͬ�����ֳ����Եĸı䡣
��
����3. 2 ��ͬ�����ݶ�����Ⱥ�ռ�ֲ����ݺ�������
������ͬ�����ݶ���Ⱥ��������������ֲ�����ʵ�ֲ��ıȽ���ͼ 2��ֱ�� y = x Ϊ��Ⱥ������ֲ�(����ֲ�) ����,ֱ�� y = ax + b(a,b ��Ϊ����,��ͼ 2) Ϊ�۲�ֵ��ʵ�ʷֲ����ߡ������ݺ�������,���������ֶ���������ֲ�,�����е�ɢ�㶼Ӧ����ֱ�� y = x ��; ���ع����� y = ax + bλ�� y = x �Ϸ�,����ֲ��Ⱥ�����������ڼ��зֲ�; ��λ�� y = x �·�,�����ֲ��Ⱥ�����������ھ����Էֲ�����нϵ͵Ŀռ������ԡ�
����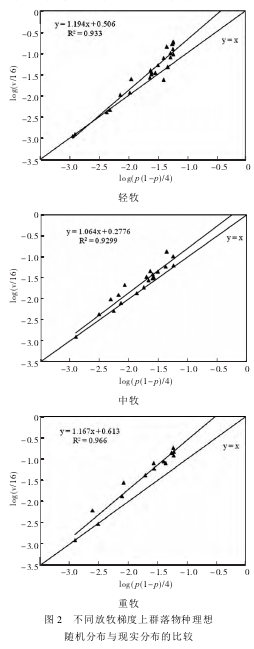
������ͼ2 ���Կ���,3 �������ݶ��ϵ�ʵ�ʷֲ����߾�λ����������ֲ����� y = x �Ϸ�,������3 ��Ⱥ��������ʵ�ʷֲ��Ͼ������,�ֲ����н�ǿ�Ŀռ������ԡ�����ȷ�����,һЩ������������ֲ�������,˵����Щ��������Ȼ��ֲ���������������ֲ������ơ����ŷ���ǿ�ȵ�����,���ֵķֲ�Խƫ������ֲ� y = x ����,˵���ڷ���ѹ������������������ֲ�������������
����3. 3 ��ͬ�����ݶ������ֳ���Ƶ����Ⱥ��ռ�������ָ���ıȽ�
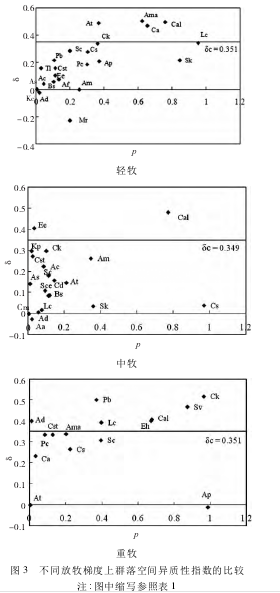
������ͬ�����ݶ���Ⱥ��ռ�������ָ���ıȽ���ͼ 3��
�������ֳ���Ƶ�� p �Ϳռ�������ָ�� δ ��ͬ�����˸�������Ⱥ�����������Էֲ�����������ú��ס�����Ƶ��Խ���Ҿ��нϸߵĿռ���������,��Խ���������Ⱥ������Ŀռ�������,��֮,����Ƶ�ȵ���������ָ���͵����ֽ�ʹȺ�������������½�����ͼ 3 ���Կ���,��ȷ����ݶȺ��ضȷ����ݶ��ϵ�Ⱥ��ռ�������ָ����ͬ,�жȷ����ݶ���Ⱥ��ռ���������͡�
������ȷ����ݶ��ϳ���Ƶ�ʺ�������ָ�����ϸߵ������� Ama��Ca��Cal��Lc; ����Ƶ����Խϵ�,������ָ���ߵ������� At��Ck; �� Sk ����Ƶ�ʽϸߵ�������ָ��ȴ����,��˵������ֲ��������ʱ���нϺõ�����ֲ��ԡ��жȷ����ݶ��ϳ���Ƶ�ʸߡ������Ըߵ�����ֻ�� Cal; ���� Ee ����Ƶ�ȵ͵������Ը�,���������ִ�Χ����,����Ⱥ������Ⱥ������Ŀռ�������ָ�����ضȷ����ݶ��Ͽռ������Ը���Ⱥ������ռ������Ե����ֹ��� 7 ��,�ֱ��� Ck��Pb��Sv��Cal��Lc��Eh �� Ad,��Щ������Ӱ��Ⱥ��ռ�������ָ����������Ҫ���á�
����3. 4 ��ͬ�����ݶ�������ƫ���ݺ����������̶߳ȵıȽ�
�������ֳ���Ƶ����ƫ���ݺ�������ֱ�̶߳ȵĹ�ϵ��ͼ 4��
����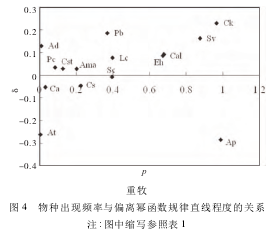
������ͼ 4 ��֪,�����ݶ��� ε≥0 �����ְ���Lc��At��Ama �� Cal �� 13 ��ֲ��,ռȺ������������һ������; �жȷ����ݶ��� ε≥0 �������� Ck��Cal��Ee �� Kp �� 9 ���������; �ضȷ����ݶ��� ε≥0 �������� Ck��Pb��Sv �� Ad �� 11 ��������ɡ�
����˵����Щ�����Դ��ڸ�Ⱥ������Ŀռ������Ե�����,�����Ⱥ����������������˹ؼ�������,����Щ���ֵij���Ƶ��Խ��Խ����������Ⱥ�������ռ������ԡ���֮,ε <0 �������ڳ���Ƶ�ʽϸ�ʱ,���ʹȺ������Ŀռ��������½����� 3 �������ݶ��ϵ���������,�����������ڵĹ㷺�ԡ������������Ե�����ռ�������ƫ��,����Ⱥ���г���һ������ֲ�������̬��,���,�ڿռ��Ͻ�����Ⱥ������Ŀռ�������ָ��,�ھ����ռ����γ������õĽṹ,�Ӷ��ܺõ�ά�ֲ�������Ⱥ��������Ե��ȶ���������
����3. 5 ��ͬ�����ݶ���Ⱥ���������ۺϱȽ�
������ͬ����ǿ����Ⱥ��������������ԵıȽϼ��� 2��
����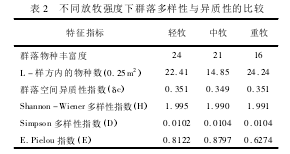
�����ɱ� 2 ��ͳ�����ݿ�֪,���ŷ���ǿ�ȵ���ǿ,Ⱥ�����ַḻ������; L �������г����������ı仯����Ϊ���� > ���� > ����; Ⱥ��ռ�������ָ���ı仯����Ϊ���� = ���� > ����;Shannon - Wiener ������ָ���� Simpson ������ָ���ڲ�ͬ�����ݶ��Ͼ�û�б��ֳ����ԵIJ����Ա仯; �� E. Pielou ���ȶ�ָ���ı仯����Ϊ����> ���� > ������˵�������ѡ���Բ�ʳ��ʹ����������������,ʹȺ��ľ�����ָ���ı�,�����ı�Ⱥ��Ŀռ������ԡ�
����4�� �� ��
���������ݺ�������Է�����������Ⱥ�Ŀռ������ԵĶ�������,�ܹ���Ϊ�ۡ�ȷ�ط�ӳֲ����Ⱥ�Ŀռ�������������ͨ�������ֳ���Ƶ����ռ�ֲ������Թ�ϵ���о�,����ֱ�۵Ŀ���Ⱥ������Ŀռ�ֲ�����,�������Ⱥ��ĸ����ֲ���ֵĿռ������Գ̶�Ҳ������������,�÷�����Ӧ�ý����Ժ��һ���Ʋ�Ⱥ��ĸǶȡ���������ռ������ԵĹ�ϵ�춨���õĻ�����
�����ݺ�������Ĺ㷺Ӧ��,�ȼ�����ֲ��������Ͷ��������������Ͳ���,���ֲ�������鷨�Բ�ԭ���������ƻ�,ͬʱ�����Ա���Ƕȵ��鷨�вⶨȺ������ʱ��������Ϊ��
����������ı���Ⱥ��ṹ,��Ӱ���ԭȺ�����ֿռ�ֲ������Ե���Ҫ��Դ�����ӡ��ڻָ����������,��Ⱥ��̶ֳȺ�Ⱥ������ۼ��̶ȵı仯����Ⱥ��̬��Ⱥ��ṹ�Ŀռ����������Ӧ,��Ⱥ�ۼ��ֲ�����ijЩ��̬���ӿռ������ԵĽ��,����Ⱥ�����������Ӱ��Ⱥ��ռ������Ե���Ҫ���֡���ͬ�����ݶ��ϲݵ�ֲ����֡����ֽṹ��ռ��������������,������������������,�ݺ������ɵĻع�ֱ��λ������ֲ�ֱ�� y = x ���Ϸ�,������ͬ�����ݶ��ϲ�ԭֲ��Ⱥ�������ϱ�����ֲ�����ֵ���и��ߵĿռ�������,�����ּ�Ⱥ�ֲ�,���־ۼ��ֲ����������ܡ��¿������ݳ˷��������������ɲ�ԭȺ��ṹ�������о������һ��,���Զ��ȶԲ�ͬ���εIJ�ԭȺ����ݺ��������о����һ�¡�
������ 0. 25 m ×0. 25 m ��С���������Ϊ��Դ�ݶ�״̬�ĺ���ָ��,�������ֵij���Ƶ��,���Զ���С�߶���Ⱥ��Ŀռ������ԡ�ֲ�����������ֵĽṹ��������Ҫ�����ڽ�Ⱥ�ֵ�����ͷ���ָʾ�ֵĸ�����,�ڲ�ͬ������ʽ��ֲ��Ⱥ�����ͼ�������Ⱥ���еĹ���λ�����÷������Ըı䡣�ڱ��о���,�ɽ�� 2. 1 ��֪,���ŷ���ѹ���ij�������,Ⱥ��������������½�����,���ڸý����,��ȡ��жȺ��ضȷ�����Ⱥ��������ָ��û�б��ֳ���Ϊ���Եı仯����,˵���ڷ���ѹ���ĸ�����,Ⱥ����ɽṹ�䷢�����Ըı�,��Ⱥ��Ŀռ�������ָ��ȴ����Ϊ���� = ���� > ����,����δ��֮�������Եĸı�,Ҳ����˵����Ⱥ��ռ�������ָ����ָ�겢��������������,����ȡ����������Ⱥ���еĹ�������,�������ֵij���Ƶ�ʼ���ռ�������ָ����ͬ������ǿ�ҵķ���������������̬�ṹ�ռ������Ե���Ҫ����,ͬʱҲ������Ⱥ���ڻ����ݶ���ԭ�пռ�ֲ���ֵ���Ҫ���ء�
����5�� �� ��
����(1) ������ֲ�����ֶ����Ա仯���������ء����ڶ���ļ�̤����ʳ�����ѻ���������,ʹ����������ʧ,�������������,�����������,������������־ͻ�������������,ѡ���зֲ�,�ı�Ⱥ����ɽṹ,�Ӷ��ı�Ⱥ��������ָ�������,����˵����������Ⱥ�����ֲַ��Ŀռ���������ҪӰ�����ء�
����(2) ���ŷ���ѹ���ij�������,���Ⱥ������������½�����,�ݺ������ɵĻع�ֱ��λ������ֲ�ֱ�� y = x ���Ϸ�,������ͬ�����ݶ��ϲ�ԭֲ��Ⱥ�������ϱ�����ֲ�����ֵ���и��ߵĿռ�������,�����ּ�Ⱥ�ֲ�; ���ֵij���Ƶ�ʼ���ռ�������ָ����ͬ������Ⱥ������Ŀռ�������ָ����
����(3) ��0. 25 m ×0. 25 m ��С���������Ϊ��Դ�ݶ�״̬�ĺ���ָ��,�������ֵij���Ƶ��,����С�߶���Ⱥ��Ŀռ�������,����ֱ�۵ط�ӳ������Ⱥ���еĹ��͵�λ,���ܸ�Ϊ���ر���Ⱥ������Ŀռ�������������
���������:
����[1]����,����. ��̬ѧϵͳ�Ŀռ�������[J]. ��̬ѧ��,2003,23(2) : 346 -352.
����[2]����Ȩ,�����,�����. ������������Ҫ���ֵĿռ�������������ȽϵĶ����о�[J]. ֲ����̬ѧ��,2000,24(6) : 718 - 723.
����[3]����. ��ͬ�ݵ�ֲ��Ⱥ��Ľṹ�����о�[D]. ����:����ũ�ֿƼ���ѧ,2012.
����[4]������,�¿�,������,��. ���������²�ͬ���Ͳݵ�Ⱥ��ṹ����[J]. ��̬ѧ��,2013,33(2) : 425 -434.
����[5]TAYLOR L R. Aggregation,variance and the mean[J].Nature,1991,189 : 234 - 240.
����[6]SHIYOMI M,TAKAHASHI S,YOSHIMURA T,et al.Spatial heterogeneity in a grassland community: use ofpower law[J]. Ecological Research,2001,16(3) : 487- 495.
����[7]����,Ѧ�,����,��. ���ױ�����ԭɳĮ�����ض�̬�仯�����о�[J]. �й�ɳĮ,2009,29(3) : 397 -403.
����[8]���Ƹ�,����ƽ,����,��. ���ױ�����ԭɳĮ����״����չ̬����������[J]. �й�ɳĮ,2005,25(5) : 635~ 639.
����[9]֣���Q,����,������,��. ���ױ�����ԭ��ͬ�ݵ����÷�ʽ��������������������Ĺ�ϵ[J]. ��̬ѧ��,2008,28(11) : 5392 -5400.
����[10]�μ�����,����̩��,�¿�. ���ڷ����ݵ�ֲ���ĵ��鷽��[J]. �ݵ�ѧ��,2005,13(2) : 149 -158.
����[11]������,�μ�����,�ŷ��,��. �����ݺ�������Է����ݶ�����Ⱥ�ռ������ԵĶ�������[J]. �й�ɳĮ,2011,31(3) : 689 - 696.
����[12]����,�¿�. ��ͬ���÷�ʽ�¶�����ɲ�ԭȺ��ṹ�������о�———�ݳ˷������Ӧ��[J]. �ݵ�ѧ��,2011,19 (3) : 388 - 394.
����[13]��Զ��,����,�¿�. ��ͬ���ι�ɽ��ԭȺ��Ľṹ����[J]. ��ҵ��ѧ,2013,30(2) : 190 -195.





