
长骨的生长是一个软骨内成骨的过程,在长骨的生长板,软骨的增殖、肥大以及基质的合成都导致新软骨的形成,同时在生长板的干骺端,新生的血管和骨细胞前体进入软骨肥大细胞带和矿化或非矿化的软骨基质区,促使软骨转变为成骨。 长骨的纵向生长速度主要是由软骨的形成速度所决定的。而软骨的形成受多种内分泌和旁分泌因子的调节。有证据显示:在长骨的生长过程中,胞外钙在调节生长板的软骨细胞功能时起到重要作用。 研究发现:钙和维生素 D 不足会导致生长板的矿化受损。
研究已经证实胞外钙发挥其生理学功能是通过激活其受体———钙敏感受体 (calcium sensing receptor,CaSR)来实现的。 CaSR 是一个 G 蛋白偶联受体,表达于哺乳动物的大多数组织中, 如甲状旁腺、骨骼、乳腺、肾脏等。 也表达于长骨生长板的增殖带和肥大带。 CaSR 基因敲除的小鼠显示生长延迟,这说明 CaSR 具有促进骨骼生长的作用。 然而这种基因敲除的小鼠表型复杂,包括甲状旁腺功能亢进、高钙血症、低磷血症,可能掩盖了 CaSR 对骨骼的直接作用。为了证实 CaSR 是否调节生长板的软骨形成以及促进长骨的生长。 本研究采用骨骼移植的方法将出生后 3 d 同窝的野生型(WT)和 CaSR 基因敲除(CaSR- /-)小鼠的股骨、胫骨移植至同父母来源的 8 周龄野生型(WT)小鼠的背阔肌中。移植骨生长4 周后,取出股骨和胫骨进行 X 线和组织学分析。
1、 材料和方法
1.1 材料
选 用 C57BL/6J 出生后 3 d 同窝的 WT 和CaSR- /-小鼠以及与该小鼠同父母来源的 8 周龄 WT小鼠各 6 只(从加拿大 McGill 大学引进,饲养于南京医科大学动物中心),雌雄不限。
1.2 方法
1.2.1 动物基因型的鉴定
取出生当天的小鼠鼠尾用苯酚-氯仿-异戊醇的方法提 DNA。 然后通过 PCR 进行基因型鉴定。
CaSR 的正向 引物序列为 :5′ -TCTGTTCTCTTTAG-GTCCTGAAACA-3′ , 反 向引 物为 :5′ -TCATTGAT-GAACAGTCTTTCTCCCT-3′,Neo 的正向 引物为 :5′-TCTTGATTCCCACTTTGTGGTTCTA-3′ , 反向引物序列同 CaSR 的反向引物序列。 所有引物均由美国Invitrogen 公司合成。Taq 酶(大连宝生物有限公司)。
PCR 反应条件 :94℃ 预 变 性 4 min,94℃ 解链 30 s,55℃退火 30 s,72℃延伸 30 s。 共进行 30 个循环 。72℃延伸 10 min。 使用 Bio-Rad Mycycler PCR 扩增仪。 CaSR 的扩增产物为 221 bp,Neo 扩增产物为276 bp。 只有 CaSR 扩增条带的为野生型(WT),只有Neo 扩增条带的为纯合子(CaSR- /-),既有 CaSR 扩增条带,又有 Neo 扩增条带的为杂合子。 本试验中使用的是 WT 和 CaSR 纯合子(CaSR- /-)小鼠(图1)。
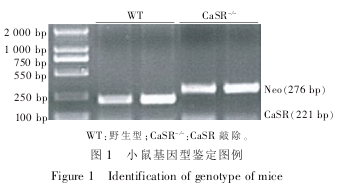
1.2.2 骨移植
取 C57BL/6J 来源的出生后 3 d 同窝的 WT 和CaSR- /-小鼠左侧的股骨和胫骨,移植至与该小鼠同父母来源的 WT 小鼠的背阔肌中。 左侧背阔肌移植WT 小鼠的股骨、 胫骨, 右侧移植 CaSR- /-小鼠的股骨、胫骨。 移植骨在背阔肌中生长 4 周。
1.2.3 取材
将 3 d 的 WT、CaSR- /-小鼠左侧的股骨、胫骨和生长 4 周的移植骨取出,放在多聚甲醛-赖氨酸-过碘酸钠固定液中固定,常规脱钙,脱水,石蜡包埋,然后切片做组织学染色。
1.2.4 X 线摄影
未脱钙的 WT 和 CaSR- /-小鼠的股骨、胫骨使用Faxitron model 805 radiographic inspection system(Faxitron Contact 公司, 德国) 进行摄片 (电压 22kV, 曝光时 间 4 min)。 底片 使用 X-Omat TL film(Eastman Kodak 公司,美国),常规方法进行显影定影。 使用 HP 扫描仪扫描分析。
1.2.5 HE 染色
石蜡切片常规脱腊水化,苏木素(Sigma 公司,美国)染色,1%盐酸酒精分色,流水冲洗返蓝,伊红(Sigma 公司,美国)复染,常规脱水透明 ,中性树胶封片。
1.3 统计学方法
所有染色均由 Leica 数码显微镜拍摄, 并用Northern Eclipse 图像分析软件进行定量分析。 数据使用 SPSS15.0 软件进行组间单因素方差分析,P≤0.05 为差异具有统计学意义。
2、 结 果
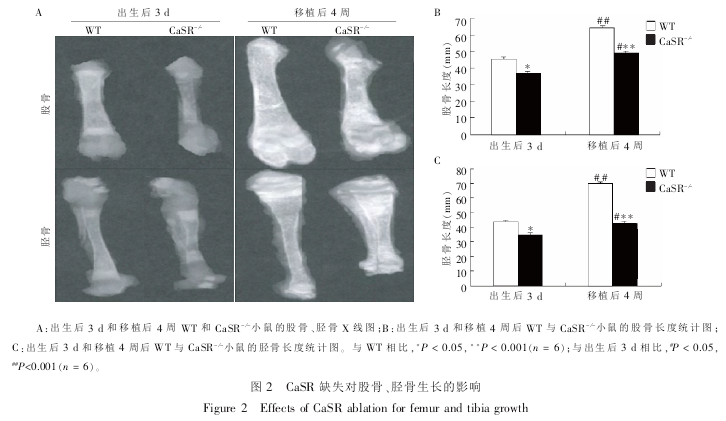
2.1 CaSR 缺失对股骨、胫骨生长的影响
为了验证 CaSR 缺失是否会引起长骨长度的改变, 通过 X 线检测了生后 3 d 以及骨移植 4 周后的WT 和 CaSR- /-小鼠的股骨和胫骨的长度。结果显示:出生后 3 d,CaSR- /-小鼠的股骨、 胫骨长度相对于WT 小鼠明显缩短 (P < 0.05)。 移植 4 周后,WT 和CaSR- /-小鼠的股骨、 胫骨相对于出生后 3 d 时均显著增长(P < 0.001,P < 0.05),但 WT 小鼠增长的幅度远远大于 CaSR- /-小鼠增长的幅度(图 2)。 这说明CaSR 具有促进长骨纵向生长的直接作用。
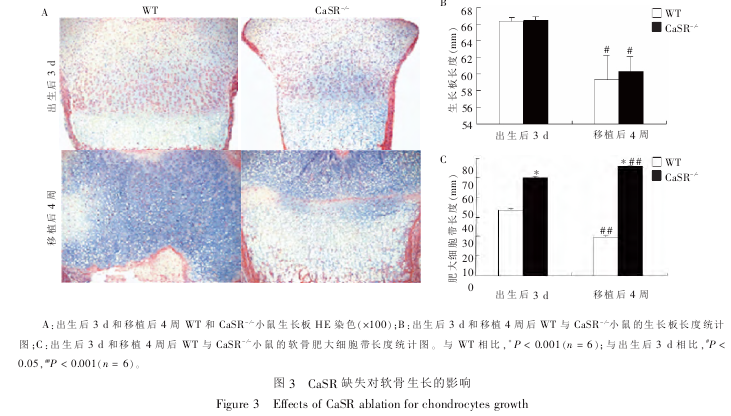
2.2 CaSR 缺失对软骨生长的影响
为了研究 CaSR 缺失是否对软骨生长产生影响,通过石蜡切片 HE 染色对股骨生长板的长度、生长板的增殖带和肥大带进行观察。结果显示:无论是在出生后 3 d 还是移植后 4 周,WT 和 CaSR- /-小鼠股骨生长板的长度两者之间没有明显差异, 但是 4周时股骨生长板相对于 3 d 时的生长板显著缩短(P< 0.05)。 这可能与 4 周时长骨的纵向生长已经减弱或停止,生长板在骨生长中的作用逐渐减弱有关。但从生长板肥大细胞带的长度可以看出: 无论是 3 d还是移植后 4 周,CaSR- /-小鼠的肥大细胞带均显著宽于 WT 小鼠(P < 0.001)。 移植 4 周后,WT 小鼠股骨肥大细胞带要显著小于出生后 3 d 时 WT 小鼠的肥大细胞带(P < 0.001);但是移植 4 周后,CaSR- /-小鼠股骨肥大细胞带比出生后 3 d 时 CaSR- /-小鼠的肥大细胞带显著增宽(P < 0.001,图 3)。
3 、讨 论
先前 Garner 等对 WT 和 CaSR- /-小鼠的骨骼表型做了详细分析, 发现敲除甲状旁腺中的 CaSR基因将导致甲状旁腺功能亢进、高钙血症、低磷血症以及骨骼中的 PTH 过量表达,而且 CaSR 基因敲除的小鼠显示出骨骼生长的延迟,其软骨发育不全,增加矿化骨的形成和骨吸收, 不能纠正 CaSR 缺失引起的骨骼表型异常。 而且,这类小鼠最主要的骨骼异常就是佝偻病。 这说明 CaSR 具有促进骨骼生长的作用。 但它并不能反映 CaSR 对骨骼生长的直接作用。 因为 CaSR 基因敲除小鼠其表型很复杂,包括:甲状旁腺功能亢进、高钙血症、低磷血症。 这些原发或继发性的作用掩盖了 CaSR 对骨骼的直接作用。 为了证实 CaSR 是否调节生长板的软骨形成以及促进长骨生长,本研究采用骨骼移植将出生后3 d 同窝的 WT 和 CaSR- /-敲除小鼠的股骨、胫骨移植至同父母来源的 8 周龄的 WT 小鼠背阔肌中。 移植骨生长 4 周后,取出股骨和胫骨进行 X 线和组织学分析。
在出生后 3 d,WT 小鼠的股骨、 胫骨长度比 CaSR- /-小鼠的股骨、胫骨长,这可能是因为 CaSR 缺失直接导致了长骨生长的延滞,也可能是由于甲状旁腺功能亢进使得甲状旁腺激素分泌过多从而抑制骨骼的生长。
为此,本研究采用骨骼移植的方法给 CaSR- /-小鼠骨骼的生长提供一个与 WT 小鼠骨骼生长相同的环境。 移植 4 周后,虽然 WT 和 CaSR- /-小鼠的长骨都有增长,但 WT 小鼠的增长幅度远远大于 CaSR- /-小鼠。 这说明 CaSR 的缺失可以直接抑制骨骼生长。
长骨的生长过程是一个软骨内成骨的过程。 那么 CaSR 缺失抑制长骨的生长是否是通过影响软骨的形成来实现呢? 为此研究了生长板的软骨细胞。
HE 染色显示无论是出生后 3 d 还是移植后 4 周,生长板的总长度在两种基因型小鼠(WT 和 CaSR- /-)之间没有明显差异,但在移植后 4 周时,两种基因型小鼠的生长板长度要比生后 3 d 时短, 这主要是与移植 4 周以后,移植骨的骨龄已趋成年化,长骨的纵向生长已停止或减弱,软骨细胞的功能减弱有关。 虽然生长板的总长度在两种基因型小鼠之间没有明显差异,但软骨肥大细胞带却有明显差异,无论是在出生后 3 d,还是在移植后 4 周,CaSR- /-小鼠的软骨肥大细胞带明显宽于 WT 小鼠;在 WT 小鼠中,移植后4 周的肥大细胞带要小于出生后 3 d, 而在 CaSR- /-小鼠中,移植 4 周后的肥大细胞带要宽于出生后 3 d。
CaSR 在生长板中的表达主要位于生长板的增殖带和肥大细胞带。 所以 CaSR 缺失以后,软骨肥大细胞带增宽,骨长度缩短,这说明 CaSR 缺失使得软骨肥大细胞向成骨细胞矿化、骨化能力降低,软骨细胞聚集在肥大细胞带,从而使肥大细胞带增宽。
为了完全了解 CaSR 对骨和软骨功能的直接作用,纠正甲旁亢是必须的。 因为 CaSR- /-新生小鼠致死率很高,从而掩盖了成年鼠中 CaSR 的骨骼作用。
最近,有两个研究小组利用了双敲模型,Tu 等利用 CaSR 与 Gcm2 双基因敲除,Kos 等利用 CaSR和 PTH 双基因敲除,成功纠正了甲旁亢以及与之伴随的高钙血症和低磷血症。在双敲模型中,小鼠骨骼没有明显异常,这说明:在骨骼分化和重建过程中,CaSR 并非是必不可少的。 但这并不能否认 CaSR对骨骼的作用。因为对于维持细胞的正常功能来说,钙的调节是最基本的,CaSR 的缺失可能导致其他代偿途径, 其中一个代偿机制就是骨细胞中 CaSR 剪切变体的表达。 这些受体存在于骨骼中,对胞外钙浓度的改变以及机械应力的改变具有反应。
而且有人报道存在另一种不同于 CaSR 的 G 蛋白偶联受体。 所有这些可能解释以上双敲小鼠骨骼表型没有改变的现象。
影响软骨内矿化和骨化的因素很多, 包括多种激素和调节因子,通过影响软骨细胞分化速度、软骨细胞间信号转换、 软骨细胞肥大以及软骨细胞所表达的特有基质类型等来协调软骨基质的钙化、 矿化和骨化过程。 这些激素和调节因子之间又存在相互作用,并通过复杂的反馈机制对靶器官产生作用。因此, 本研究只揭示 CaSR 对软骨细胞的作用主要集中于软骨细胞的肥大向矿化的过程中, 至于 CaSR对软骨细胞矿化、 骨化影响的具体机制还需进一步的研究。





