摘 要: 西藏蟾蜍 (Bufo tibetanus) 主要生活在海拔2 400~4 300 m的高海拔地区, 本研究分析了这一高原两栖物种雄性个体的鸣声特征和听觉敏感性。采用录音机和指向性话筒, 在野外记录西藏蟾蜍的广告鸣声, 使用听觉脑干反应 (ABR) 检测听觉敏感性。采用Praat声音分析软件绘制广告鸣声的波形图和频谱图, 鸣声特征参数通过Adobe Audition软件获取。广告鸣声由多个单音节鸣叫组成, 鸣声主频为 (1150±99) Hz。ABR对于刺激的响应以谷峰波形展示, 听力图结果显示, 听觉敏感区域在1.4~2.0 kHz, 但在0.6~6.0 kHz范围的听觉阈值均高于70 dB, 表明雄性西藏蟾蜍相较于其他物种听觉敏感性较差。尽管雄性西藏蟾蜍的最佳听觉敏感频率 (1.6kHz) 稍高于鸣声主频, 但其鸣声能谱结构与听觉敏感性曲线在1.0~1.4 kHz存在一定程度重叠, 符合“匹配过滤假说”。
关键词: 鸣声特征; 听觉脑干反应; 匹配过滤假说; 西藏蟾蜍;
Abstract: The Tibetan Plateau Toad (Bufo tibetanus) lives in the plateau regions at 2 400-4 300 m above sea level. This is the first report to analyze the call characteristics and auditory sensitivity of male toad. Calls of 6 individuals were recorded by directional microphone connected to a digital audio recorder. Auditory sensitivity of 13 males was measured by auditory brainstem response (ABR) . The waveform and spectrogram of advertisement calls were drawn using Praat software (Fig. 1) . The acoustic properties were obtained using Adobe Audition software. The advertisement call of male toad consists of a single note with a dominant frequency of 1 150 ± 99 Hz. ABR to tone pips were characterized by valley-peak waveforms (Fig. 2) . The audiogram shows that the most sensitive frequency region is 1.4-2.0 kHz, but the ABR thresholds are above 70 dB across 0.6-6.0 kHz which means that this toad has a poor auditory sensitivity compared with other species (Fig. 3) . The power spectra of the male call overlap the audiogram at 1.0-1.4 kHz, which supported the matched filter hypothesis, although the best auditory sensitive frequency (1.6 kHz) is slightly higher than the dominant frequency of the male call.
Keyword: Call characteristics; Auditory brainstem response; The matched filter hypothesis; Bufo tibetanus;
声音通讯是无尾两栖类个体间最主要的交流手段, 与在繁殖季节的雄性竞争和吸引雌性等行为密切相关 (Wells 1977, Gerhardt et al.2002) 。在多数情况下由雄性个体发出鸣叫, 声音信号在传递过程中携带多种信息, 包括信号发出者的身体大小、位置信息以及是否有繁殖场所等 (Cui et al.2012, Zhu et al.2017) 。
听觉敏感性反映了个体对声音信号的接收能力, 无尾两栖类的听觉敏感性主要由其内耳的两栖乳突 (amphibian papilla) 和基底乳突 (basilar papilla) 决定。其中, 两栖乳突决定低、中频听觉敏感性, 基底乳突决定高频听觉敏感性 (Smotherman et al.2000) 。听觉脑干反应 (auditory brainstem response, ABR) 能很好地估计听力图的形状, 因此是研究听觉敏感性和听觉系统功能的重要手段, 已大量应用于包括无尾两栖类在内的多个脊椎动物类群中 (Corwin et al.1982, Womack et al.2017) 。ABR是一种听觉激发电位反应, 它表征的是听觉神经和脑干神经元同步活动的总输出, 其波形由一系列典型的正负变量表征, 根据这些变量的有无来确定听觉阈值 (Stapells et al.1997, Brittan-Powell et al.2010) 。

“匹配过滤假说” (the matched filter hypothesis) 认为, 听觉敏感性与鸣声频率相匹配, 可以过滤鸣声频率以外的其他频率信号。神经过滤机制与鸣声信号特性的一致性可以有效过滤噪声干扰, 使信号传递效率最大化 (Capranica et al.1983) 。“匹配过滤假说”已在多个物种中得到支持, 包括两栖类 (Zhu et al.2017) 、爬行类 (Chen et al.2016) 以及鸟类 (Dooling et al.1971) 等。大多数无尾两栖类的听觉敏感性与种内声音信号相匹配, 以提高声音信号的信噪比 (Feng et al.2007) 。但是, 也存在不符合“匹配过滤假说”的例子, 小湍蛙 (Amolops torrentis) 的广告鸣叫主频 (4.3kHz) 与最佳听力范围 (1.6~2.0 kHz) 完全不匹配 (Zhao et al.2017) 。
西藏蟾蜍 (Bufo tibetanus) 主要分布于青海、西藏、四川及云南等海拔在2 400~4 300 m的高原草地或山区, 主要在夜间外出活动 (Fei et al.2016) 。国内已有关于西藏蟾蜍皮肤组织学结构 (高凤娟等2016) 及系统地理学研究 (Zhan et al.2011) , 但没有声音通讯相关的研究。受青藏高原低温、低氧等极端环境的影响, 西藏蟾蜍的声音通讯可能存在适应性变化。本研究通过野外鸣声录制及听觉敏感性检测等手段以了解西藏蟾蜍这一高海拔分布物种声音通讯的相关情况。为探讨高原蛙类声音通讯系统的高原适应提供研究基础。
1、 材料与方法
1.1、 研究地点和研究对象
2018年5月, 在康定市塔公乡塔公草原附近 (30°18′37.5″N, 101°30′32.7″E) 录制成体雄性西藏蟾蜍的鸣声。录音期间, 环境温度 (6.1±2.2) ℃, 相对湿度56.8%±6.5%。听觉脑干反应 (ABR) 电生理实验在将活体样本带回实验室后一周内完成, 共检测13个雄性样本, 实验期间室内温度为 (23.7±1.1) ℃。
1.2 、声音录制
野外研究期间, 晚上21至23时在西藏蟾蜍栖息水塘边使用指向性麦克风 (Sennheiser ME66 with K6 power module) 连接录音机 (Marantz PMD 661, 16 bit, 44.1 kHz) 进行录音, 麦克风距离蟾蜍1 m左右。录音时采用单声道模式, 录音文件以wav格式保存, 每段录音时长约为4 min, 包含同一个体的多次鸣叫。
1.3、 ABR检测
ABR检测在一个小型铁质隔音屏蔽箱 (长0.5 m×宽0.5 m×高0.5 m) 中进行, 实验动物均使用0.2%MS-222 (鱼安定, tricaine methane sulfonate, 北京格林恒兴生物科技有限公司) 溶液进行轻微麻醉, 直至动物对夹趾没有明显反应, 该过程一般耗时2~4 min。测量时实验对象保持自然姿势且头部垂直朝向播放刺激的音箱 (SME-AFS, Saul Mineroff Electronics, Elmont, NY) , 3根27号皮下针状电极 (Rochester Electro-Medical, Inc., FL, USA) 分别植入头顶、鼓膜和对侧前肢的皮下。电极与信号放大器 (PA4&RA4, 20×gain) 连接, 信号放大器通过光纤与数字信号处理器RM2 (Tucker-Davis Technologies, Gainesville, USA) 连接。RM2通过USB接口与电脑连接, 使用Open ABR软件操作。通过便携式音箱 (SME-AFS;Saul Mineroff Electronic Inc, USA) 播放两种声音刺激 (纯短音tone pips和短声clicks) , 其中tone pips用来测量在每个频率的听觉阈值, clicks作为对呈现生物学信号反应的验证 (作为阳性对照) 。实验时音箱与蟾蜍吻端的距离约为10 cm, 此外, tone pips刺激的范围为0.6~6.0 kHz (0.6、0.8、1.0、1.2、1.4、1.6、1.8、2.0、2.2、2.4、2.6、2.8、3.0、3.2、3.4、3.6、3.8、4.0、5.0、6.0 k Hz) , 每个刺激频率都测量9个声压梯度, 梯度间隔设置为5 dB, 由于预实验显示西藏蟾蜍听觉阈值较高, 选择的梯度范围为50~90 dB。刺激包括1 ms的上升/下降时间和3 ms的平台时间, 取样率为24414 Hz。每次测量结果由400次重复刺激的平均值产生。在每个刺激频率下均进行2次重复的ABR测量。
ABR实验前, 用G.R.A.S.46 BE 1/4英寸的麦克风 (G.R.A.S.Sound&Vibration, Denmark) 对0.6~10.0 kHz的声音刺激声压级进行校准, 校准时麦克风前端位于实验时蟾蜍头部的大概位置。
1.4、 数据统计和分析
采用Praat声音分析软件绘制西藏蟾蜍广告鸣声的波形图和频谱图, 使用Adobe Audition 3.0 (Adobe USA) 和Praat软件统计广告鸣声的主频 (dominant frequency) 、谐波数 (harmonic number) 、音节时长 (note duration) 。每只蟾蜍随机取4~6个广告鸣声, 计算鸣声主频、谐波数及音节时长的平均值作为该个体的广告鸣声特征数据, 然后对所有个体鸣声数据再次求平均。
ABR阈值的获取方法参照其他使用ABR检测无尾两栖类听觉敏感性的研究 (Zhang et al.2012, Cui et al.2017) , 即每个频率下有明显反应电位的最低声压值被视为该频率的听觉阈值, 统计时将两次测量的最低阈值作为对应频率的数值。在特定频率下, ABR阈值越低表示在该频率下听觉敏感性越好。本研究使用13个雄性个体的ABR数据, 以代表雄性西藏蟾蜍整体的听觉敏感性。ABR阈值和形态参数使用SigmaPlot 11软件 (Systat Software Inc., San Jose, USA) 进行统计分析。数据均以平均值±标准差 (Mean±SD) 表示。
2、 结果
2.1、 鸣声结构及参数
在野外共获得6只西藏蟾蜍的27个广告鸣声。图1展示了西藏蟾蜍广告鸣声的波形图 (图1a) 及频谱图 (图1b) 。西藏蟾蜍的鸣声比较单一, 广告鸣声均为短促的单音节。每个音节包含多个频率段并且由多个谐波 (16±3) 组成, 音节时长 (80.67±13.89) ms。能谱分析结果显示, 西藏蟾蜍广告鸣声主频为 (1 150±99) Hz。
2.2、 ABR波形图
ABR波形图由一系列发生于声音刺激后10 ms以内的波组成, 图2a展示了雄性西藏蟾蜍在1.6 kHz刺激下典型的ABR波形图。在图2a中能看到明显的谷峰波形结构, 但波形的振幅随着声压级的下降而减小。图2b展示了将ABR波形图部分放大后的结果, 反应波在60 dB消失, 只能观察到由噪音产生的微小杂波, 在该波形图下雄性西藏蟾蜍在1.6 kHz下的听觉阈值为65 dB。
图1 西藏蟾蜍的波形图 (a) 及频谱图 (b)
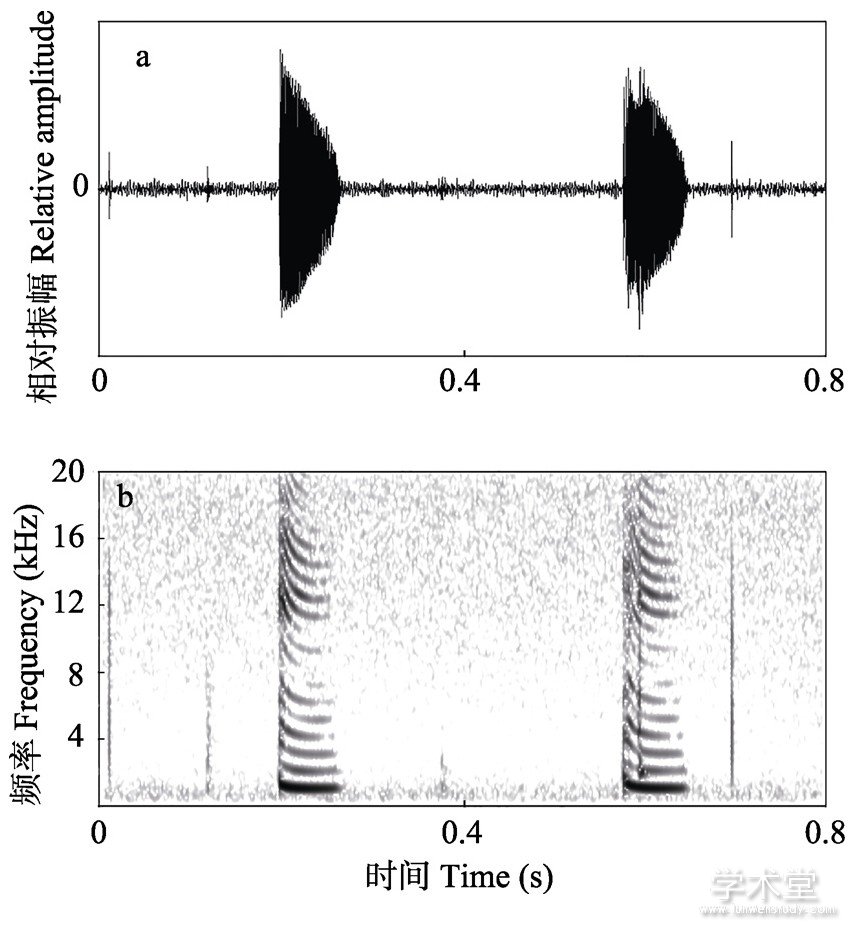
Fig.1 Amplitude-modulated waveform (a) and spectrogram (b) of Bufo tibetanus
图2 1.6 kHz频率下雄性西藏蟾蜍ABR波形图
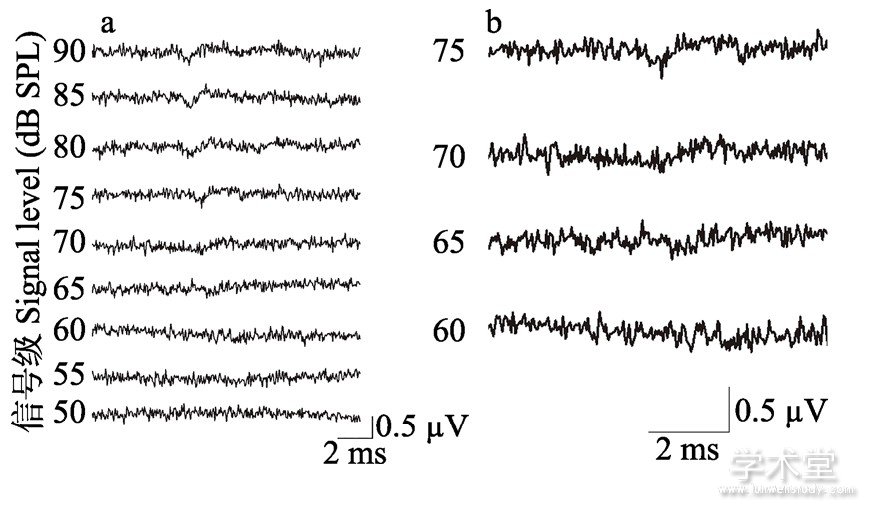
Fig.2 Representative ABR waveforms of male Bufo tibetanus in response to 1.6 kHz tone bursts
a.ABR波形图;b.放大图。a.ABR waveform;b.Enlarged view.横向箭头表示在该ABR波形图中观察到的听觉阈值。
The right-pointing arrowheads depict the visually detected threshold in the ABR waveform.
2.3、 听觉敏感性曲线及鸣声能量谱
各频率ABR阈值是在该频率下能检测到的最低刺激强度, 多个频率下的ABR阈值组成该物种的听觉敏感性曲线。本次研究共对13个雄性样本进行ABR检测。雄性西藏蟾蜍在0.6~6.0 kHz频率下的听觉敏感曲线见图3, 其听觉阈值相对较高, 整体均在70 dB以上。在1.4~2.0 kHz存在一个听觉相对敏感频段 (约75 dB) , 2.0 kHz以上, 听觉阈值呈上升趋势。图3阴影部分展示了西藏蟾蜍广告鸣声的能量谱, 西藏蟾蜍的鸣声能量主要分布在1.0~1.6 kHz。整体而言, 雄性西藏蟾蜍广告鸣声能谱结构与听觉敏感性曲线在1.0~1.4 kHz存在一定程度重叠, 但听觉敏感频率 (1.4~2.0 kHz) 较鸣声主频 (1.2 kHz) 稍高一些。
图3 雄性西藏蟾蜍听觉敏感曲线和鸣声能量谱
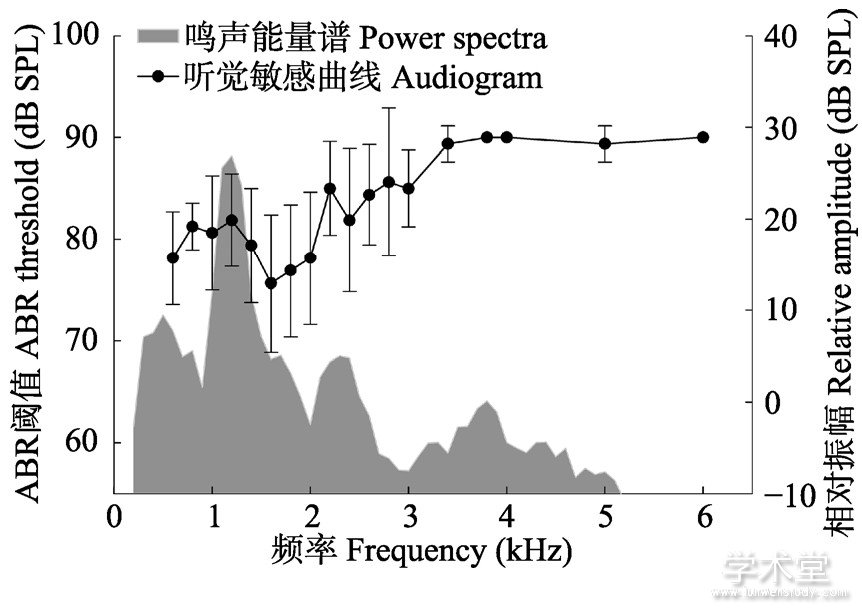
Fig.3 The audiogram and power spectra of call of male Bufo tibetanus
听觉敏感性曲线纵坐标以ABR阈值表示, 鸣声能量谱纵坐标以相对振幅表示。
The ordinate of the audiogram is represented by the ABR threshold, and the ordinate of the power spectra is represented by the relative amplitude.
3、 讨论
本研究首次分析了高原两栖物种西藏蟾蜍雄性个体的鸣声特征和听觉敏感性。西藏蟾蜍的广告鸣声由多个单音节的鸣声组成, 鸣声较为短促并包括多个谐波, 鸣声能量主要分布在1.0~1.6 kHz。雄性西藏蟾蜍ABR结果显示, 其听觉敏感曲线呈“V”字形, 存在一个听觉敏感区域 (1.4~2.0 kHz) , 最佳听觉频率为1.6 kHz, 且各个频率下的听觉阈值均处于70 dB以上。
多数无尾两栖类的ABR检测结果显示, 它们在低频和高频区域都存在一个听觉敏感区域, 最低的听觉阈值大多在45 dB至65 dB (Cu et al.2017, Womack et al.2017, Taylor et al2018) , 本研究中雄性西藏蟾蜍的听觉阈值在70 dB以上, 表明其整体听觉敏感性较差, 听力明显弱于其他无尾两栖类物种。另外, 温度对ABR敏感性具有一定影响, 在不同温度下对小湍蛙的ABR检测结果显示, 高温下的ABR阈值更低 (未发表数据) , 因此在高原低温环境下, 雄性西藏蟾蜍的听觉阈值可能更高。雄性西藏蟾蜍存在一个听觉敏感区域, 对低频敏感性的缺失可能是适应环境的结果。由于高原上刮风等原因造成的自然噪声 (<1 kHz) 经常存在, 而噪声是干扰声音传递最主要的非生物因素 (Feng et al.2007) , 雄性西藏蟾蜍的听觉敏感区域主要在1 kHz以上, 可能与此有关。部分无尾两栖类的听觉敏感性存在明显的雌雄差异 (Zhu et al.2017, Yang et al.2018) , 本研究仅包含雄性个体, 有关于西藏蟾蜍听觉敏感性的雌雄差异有待进一步研究。
“匹配过滤假说”已在多个无尾两栖类物种中得到证明, 尤其是在背条螳臂树蛙 (Chiromantis doriae) 和侧条费树蛙 (Feihyla vittata) 这两种同域分布物种的比较研究中, 两个物种均表现出种内的听觉敏感性与鸣声主频更为匹配, 以减少种间的繁殖干扰 (Yang et al.2018) 。雄性西藏蟾蜍广告鸣声能谱结构与听觉敏感性曲线在1.0~1.4 kHz存在一定程度重叠, 但最佳听觉敏感频率 (1.6 kHz) 稍高于鸣声主频 (约1.2 kHz) , 存在错位现象。错位的情况在锯腿原指树蛙 (Kurixalus odontotarsus) 中也有发现, 其鸣声主频位于两个听觉敏感频率之间, 以同时刺激两栖乳突和基底乳突 (Zhu et al.2017) 。与锯腿原指树蛙相比, 西藏蟾蜍雄性个体最佳听觉敏感频率偏高可能是更好地匹配广告鸣声多个谐波的结果, 多个谐波均有较高的鸣声能量分布, 其仍然符合“匹配过滤假说”。
西藏蟾蜍主要分布在高海拔地区, 已有研究显示, 其皮肤结构明显表现出与强紫外辐射和缺氧环境相适应的特点 (高凤娟等2016) , 西藏蟾蜍的听觉系统退化是否也是高原环境适应的一种表现?首先, 声音通讯中鸣叫行为是十分耗能的过程 (Wells et al.2007) , 高原低温低氧多风的极端环境对西藏蟾蜍的声音通讯过程是个极大的挑战, 这是西藏蟾蜍在声音通讯上产生适应性进化的可能原因。此外, 西藏蟾蜍虽然有鼓膜, 但其听力明显弱于大部分无尾两栖类, 而除西藏蟾蜍外, 部分高原蛙类也表现出明显的外周听觉系统退化的现象, 如高山倭蛙 (Nanorana parkeri) 鼓膜和耳柱骨消失 (费梁等2012) , 这将严重影响声音信号的接收和识别。基于以上两点我们推测, 西藏蟾蜍外周听力较差可能与对高原环境的适应有关, 然而这一推测仍需更多证据支持。关于无尾两栖类的声音通讯如何适应高原环境这一问题及其具体机制仍待更深入的研究。
参考文献
[]Brittan-Powell E F, Christensen-Dalsgaard J, Tang Y Z, et al.2010.The auditory brainstem response in two lizard species.The Journal of the Acoustical Society of America, 128 (2) :787-794.
[]Capranica R R, Moffat A J M.1983.Neurobehavioral correlates of sound communication in anurans//Ewert J P, Capranica R R, Ingle D J.Advances in Vertebrate Neuroethology.Boston:Springer, 701-730.
[]Chen J F, Jono T, Cui J G, et al.2016.The acoustic properties of low intensity vocalizations match hearing sensitivity in the webbed-toed gecko, Gekko subpalmatus.PLoS One, 11 (1) :e0146677.
[]Corwin J T, Bullock T H, Schweitzer J.1982.The auditory brain stem response in five vertebrate classes.Electroencephalography and Clinical Neurophysiology, 54 (6) :629-641.
[]Cui J G, Tang Y Z, Narins P M.2012.Real estate ads in Emei music frog vocalizations:female preference for calls emanating from burrows.Biology Letters, 8 (3) :337-340.
[]Cui J G, Zhu B C, Fang G Z, et al.2017.Effect of the level of anesthesia on the auditory brainstem response in the Emei music frog (Babina daunchina) .PLoS One, 12 (1) :e0169449.
[]Dooling R J, Mulligan J A, Miller J D.1971.Auditory sensitivity and song spectrum of the common canary (Serinus canarius) .The Journal of the Acoustical Society of America, 50 (2B) :700-709.
[]Fei L, Ye C Y (Chengdu Institute of Biology, Chinese Academy of Science) .2016.Amphibians of China (I) .Beijing, P.R.China:Science Press, 757-759.
[]Feng A S, Schul J.2007.Sound processing in real-world environments//Narins P M, Feng A S, Fay R R.et al.Hearing and Sound Communication in Amphibians.New York:Springer, 323-350.
[]Gerhardt H C, Huber F.2002.Acoustic Communication in Insects and Anurans:Common Problems and Diverse Solutions.2nd ed.Chicago:University of Chicago Press.
[]Smotherman M S, Narins P M.2000.Hair cells, hearing and hopping:a field guide to hair cell physiology in the frog.Journal of Experimental Biology, 203 (15) :2237-2246.
[]Stapells D R, Oates P.1997.Estimation of the pure-tone audiogram by the auditory brainstem response:a review.Audiology and Neurotology, 2 (5) :257-280.
[]Taylor R C, Akre K, Wilczynski W, et al.2018.Behavioral and neural auditory thresholds in a frog.Current Zoology, zoy089, doi:10.1093/cz/zoy089.
[]Wells K D.1977.The social behaviour of anuran amphibians.Animal Behaviour, 25 (3) :666-693.
[]Wells K D, Schwartz J J.2007.The behavioral ecology of anuran communication//Narins P M, Feng A S, Fay R R.et al.Hearing and Sound Communication in Amphibians.New York:Springer, 44-86.
[]Womack M C, Christensen-Dalsgaard J, Coloma L A, et al.2017.Earless toads sense low frequencies but miss the high notes.Proceedings of the Royal Society B, 284 (1864) :20171670.
[]Yang Y, Zhu B C, Wang J C, et al.2018.A test of the matched filter hypothesis in two sympatric frogs, Chiromantis doriae and Feihyla vittata.Bioacoustics, 28 (5) , doi:10.1080/09524622.2018.1482786.
[]Zhan A B, Fu J Z.2011.Past and present:phylogeography of the Bufo gargarizans species complex inferred from multi-loci allele sequence and frequency data.Molecular Phylogenetics and Evolution, 61 (1) :136-148.
[]Zhao L H, Wang J C, Yang Y, et al.2017.An exception to the matched filter hypothesis:a mismatch of male call frequency and female best hearing frequency in a torrent frog.Ecology and Evolution, 7 (1) :419-428.
[]Zhu B C, Wang J C, Brauth S E, et al.2017.The spectral structure of vocalizations match hearing sensitivity but imprecisely in Philautus odontotarsus.Bioacoustics, 26 (2) :121-134.
[]费梁, 叶昌媛, 江建平.2012.中国两栖动物及其分布彩色图鉴.成都:四川科学技术出版社.
[]高凤娟, 李丕鹏, 王译唱, 等.2016.西藏蟾蜍皮肤的组织学观察.四川动物, 35 (3) :444-451.





