摘 要: 环状核糖核酸(circular RNAs,CircRNAs)是一类特殊的非编码RNA分子,呈封闭环状结构,由非经典剪接方式反向剪接而形成,存在于高度分化的真核生物中。越来越多的研究表明CircRNAs在成骨分化中发生变化,本文介绍CircRNAs在组织细胞成骨分化中的作用,并对其在成骨分化过程中的作用靶点及信号通路研究现状作一综述。
关键词: 环状RNA; 成骨; 分化; 调控;
Abstract: CircRNAs are a special class of non-coding RNA(ncRNA) molecules. They have a closed circular structure and are formed by reverse splicing by non-classical splicing methods. They coexist in highly differentiated eukaryotes. More and more studies have shown that CircRNAs would change during osteogenic differentiation. This paper introduces the role of CircRNAs in tissue cell osteogenic differentiation, and reviews the current research status of the targets and signaling pathways of CircRNAs during osteogenic differentiation.
Keyword: CircRNAs; Osteogenesis; Differentiation; Regulation;

环状核糖核酸(circular RNAs,Circ RNAs)首次在类病毒中被发现[1],被认为是异常剪接的副产品。Circ RNAs通过5’端和3’端的反向剪接,形成环状RNA结构[2],这种独特的环状结构可以抵抗核糖核酸酶的消化,并且相对稳定,半衰期可以超过48小时[3],随着高通量测序技术的发展,在众多的细胞系和不同的物种中已经有效地识别出了大量的Circ RNAs[4,5],大量证据表明它们在器官和疾病发育过程中的基因调控中扮演着不同的角色,在神经系统疾病、癌症以及干细胞的发育、干性维持及多向分化调控和调节分化的生物学活动中发挥关键作用[6,7,8,9]。其中,Circ RNAs在成骨过程中的变化提示,Circ RNAs在成骨过程中可能发挥作用[10,11,12,13]。本文介绍在不同组织细胞中Circ RNAs成骨分化的作用(图1),将Circ RNAs在成骨分化过程中生物学功能的研究现状作一综述。
图1 Circ RNAs在成骨分化中的作用
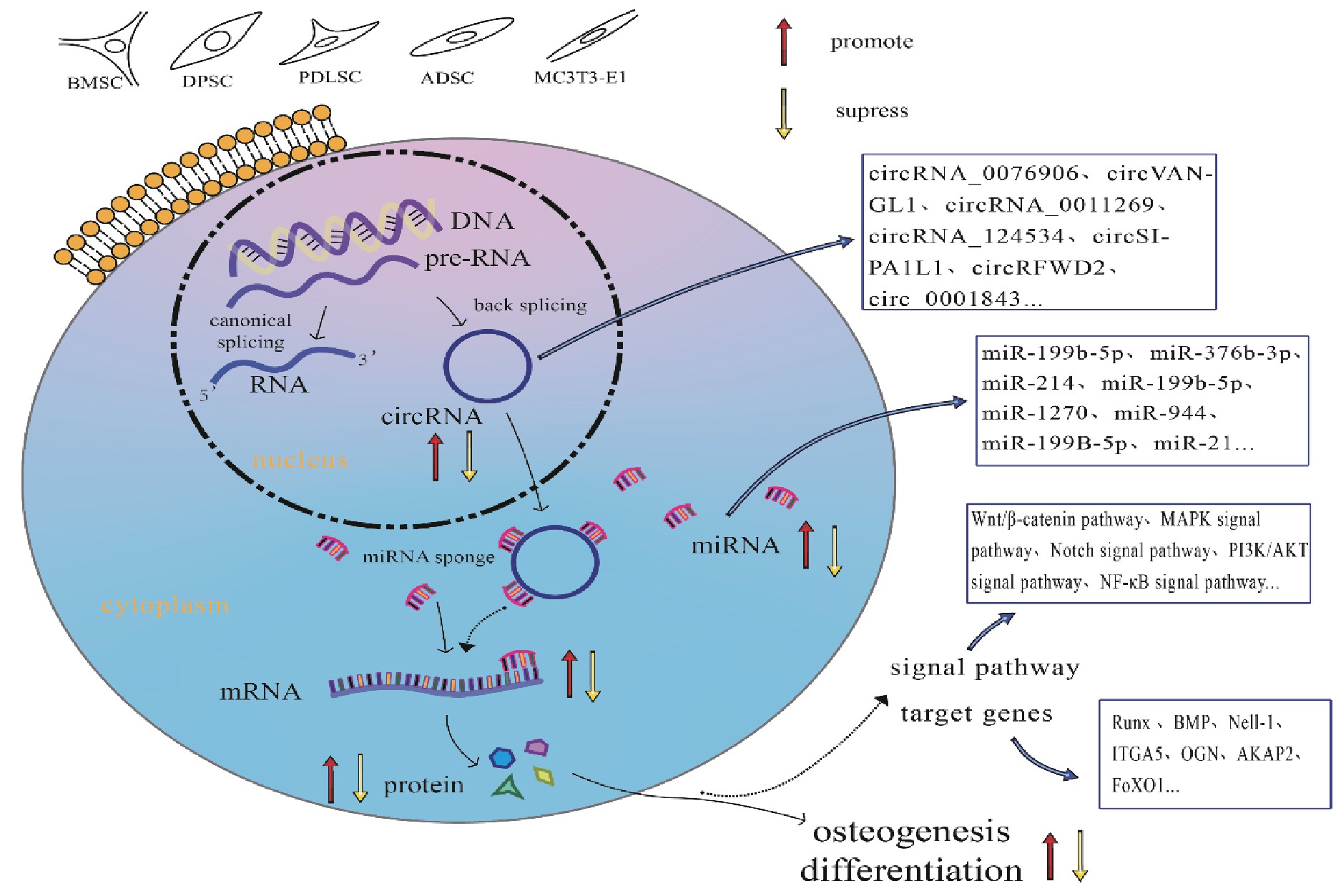
Figure 1.The role of Circ RNAs in osteogenic differentiation
1 、Circ RNAs的特点
Circ RNAs是一类新的非编码RNA,是在转录过程中RNA转录片段端到端连接而产生的闭环RNA。根据基因组起源,Circ RNAs分为三类:仅由内含子组成的内含子Circ RNAs(Ci RNAs);仅由外显子组成的外显子Circ RNAs(Ecirc RNAs);由外显子和内含子共同形成的外显子-内含子Circ RNAs(EIcirc RNAs)[14]。目前普遍认为Circ RNAs的环化方式主要有套索驱动环化、内含子配对驱动环化和RNA结合蛋白驱动环化3种[3,14,15]。套索驱动环化指当m RNA前体(prem RNA)进行GU/AG剪切时,可以跨外显子进行剪切,形成含有外显子和内含子的套索中间体,然后加工形成Circ RNAs。内含子配对驱动环化指某些外显子两侧的内含子序列中含有反向互补序列,它们之间互补配对形成RNA双链,促进Circ RNAs的形成。而RNA结合蛋白驱动环化则指一些RNA结合蛋白可以结合到外显子侧翼的内含子序列上,促进Circ RNAs的形成。
因为缺少RNA酶介导的降解的自由端[16],所以与lnc RNAs和mi RNAs相比,Circ RNAs在哺乳动物细胞中具有更高的稳定性和序列保守性[17]。也有研究发现,Circ RNAs具有种属特异性、组织特异性、疾病特异性以及与发育阶段相关的特异性[18,19,20]。同时一个基因可以产生多种不同类型的Circ RNAs,甚至有些Circ RNAs表达量比其相对应的线性RNAs还要丰富。尽管有这些发现,Circ RNAs的许多特征仍不清楚。
2、 干细胞中Circ RNAs的成骨分化作用
近年来,细胞治疗特别是间充质干细胞(mesenchymal stem cells,MSCs)在骨缺损的治疗中显示出良好的应用前景。MSCs是一种多潜能干细胞,具有自我更新和多向分化的特点。它们几乎存在于所有组织中,并在组织修复和再生中发挥重要作用[21]。常用于骨组织工程的MSCs主要包括:骨髓间充质干细胞(bone marrow mesenchymal stem cells,BM-MSCs)、脂肪源性间充质干细胞(adipose-derived mesenchymal stem cells,ADSCs)、牙髓干细胞(dental pulp stem cells,DPSCs)、胚胎干细胞、脐血间充质干细胞、诱导多能干细胞等[22]。近年来许多关于非编码RNA调控成骨分化的研究主要集中在BM-MSCs、ADSCs、DPSCs、牙周韧带干细胞(periodontal ligament stem cells,PDLSCs)等细胞系上,且许多研究已表明Circ RNAs在这类细胞成骨分化过程中发挥重要作用。
2.1 、骨髓间充质干细胞
BM-MSCs是一种多能干细胞,具有高成骨潜能,被广泛用于组织工程研究及应用。BM-MSCs是成骨细胞的主要来源,可以促进缺损区骨修复[22]。BM-MSCs的成骨分化受一系列编码和非编码RNA调控[23,24,25]。目前已经有不少研究表明Circ RNAs对BM-MSCs成骨分化过程具有调控作用,例如:在骨质疏松研究中,circ RNA_0076906、circ VANGL1、circ RNA_0011269、circ RNA_0016624、circ RUNX2、circ_0076690、circ_0024097及circ_0006393在人BM-MSCs中起到促进成骨分化,抑制骨质疏松发展的作用[26,27,28,29,30,31,32,33]。circ RNA_1983在硅酸氢钙微粒诱导BM-MSCs成骨分化和骨缺损修复中具有促进作用[34]。circ DAB1、circ RNA_33287、circ RNA_0074834、circ-SLC8A1的抑制降低了BMSCs的矿化结节形成已经被证明能够促进人BM-MSCs的成骨分化[11,34,35,36]。另外有研究表明,除了正向调控作用外,Circ RNAs对BM-MSCs成骨分具有负向调控影响,例如:circ IGSF11、circ RNA_0127781可能参与抑制人BM-MSCs的成骨分化[37]。Kuang等人揭示,在糖皮质激素引起的股骨头坏死中circ USP45在BM-MSCs的成骨分化过程中起负调控作用[38]。
此外,Zhang等证实在股骨头坏死的BM-MSCs中circ RNA_0000219、circ RNA_0004588、circ RNA_0005936等Circ RNAs的表达与BM-MSCs的增殖和成骨能力呈潜在的正相关[37]。在钛表面机械研磨促进人BM-MSCs成骨分化的研究中发现circ RNA_0032600对人BM-MSCs中晚期成骨分化也有调节作用[39]。
2.2、 牙髓干细胞
DPSCs来源于牙髓组织,可从牙髓组织中获取,具有来源广泛、较高的安全性、高增殖潜能及较强的克隆能力、可自我更新性和多向分化性等特性,已成为组织工程及再生医学领域的种子细胞[40]。DPSCs的成牙本质向分化是牙体组织自我防御及修复的关键,已有研究证实circ RNA可参与调控DPSCs的成牙本质向分化过程,例如研究表明circ RNA_0015260、circ RNA_0006984在诱导成牙本质分化的人牙髓细胞中表达上调,并证明其具有促进人牙髓细胞的成牙本质分化的功能[41]。
在不同的诱导条件下,DPSCs可以分化为神经性、成骨性和肌源性细胞系[34]。来自活体研究和动物模型的实验证据表明,DPSCs具有产生适当血管化的板层骨的能力[42]。因此,越来越多学者将DPSCs用于骨再生的研究中。其中不少学者对Circ RNAs在DPSCs成骨分化过程的作用进行研究。如体内外研究发现在人DPSCs成骨分化过程中,circ RNA_0026827的表达显着增加,且体内异位骨模型研究也发现circ RNA_0026827过表达在促进异位骨形成中起重要作用[43];另外circ RNA_124534、circ SIPA1L1也被证实能促进DPSCs向成骨细胞分化[44,45]。
2.3 、脂肪源性间充质干细胞
ADSCs位于脂肪组织的基质-血管段,由于其具有来源丰富、自体来源容易等特点,逐渐成为骨再生医学研究的重点。然而,有限的成骨分化潜能和天然的向成脂细胞分化的倾向极大地阻碍了其在骨修复中的临床应用发展[44]。ADSCs的成骨分化涉及包括非编码RNA在内的多种因素的复杂调控,而且在很大程度上还没有确定[45]。部分研究已证实Circ RNAs在ADSCs成骨分化过程中具有调控作用,例如circ RNAvgl13能显着促进ADSCs的成骨分化[46];circ RFWD2和circ INO80可以与mi R-6817-5p相互作用,从而抑制ADSCs成骨作用[47]。
2.4、 牙周韧带干细胞
PDLSCs已被证明能产生典型的牙周韧带样组织,以再生牙周炎损伤的组织。此外PDLSCs具有巨大的再生能力潜力,这有助于其自我更新和多向分化,尤其是成骨方向分化[48]。例如,Li等观察到circ CDR1as在PDLSCs成骨分化过程中显着上调,并发现circ CDR1as能促进其成骨分化[49]。而Gu等人通过数据分析发现circ BANP、circ RNA_4214、circ RNA_3140和circ ITCH对PDLSCs成骨分化具有调控作用[48]。此外circ RNA_432、circ RNA_126、circ RNA_4045、circ RNA_4251、circ RNA_5331、circ RNA_3140、circ RNA_436和circ RNA_4045等Circ RNAs被检测对机械力刺激下的PDLSCs的成骨分化具有调控作用[50]。
2.5、 小鼠胚胎成骨细胞前体细胞
小鼠胚胎成骨细胞前体细胞(MC3T3-E1)是一种成骨细胞样细胞系,是研究成骨细胞增殖和分化,以探讨骨质疏松分子机制的有用模型[51]。有报道称在小鼠胚胎成骨细胞前体细胞中circ RNA_19142和circ RNA_5846与成骨细胞分化相关[12]。mm9_circ_009056能够正向诱导小鼠胚胎成骨细胞前体细胞成骨分化[52]。另外,在氧化应激诱导小鼠胚胎成骨细胞前体细胞的细胞凋亡和成骨的研究中表明,circ_0001843具有成骨促进作用[53]。而在微重力作用下的MC3T3-E1细胞成骨过程circ_010383、circ_014154和circ_014977的差异表达较明显,且生物信息学分析发现,circ_014154靶向mi R-145a-5p和let-7a-5p,另外GO和KEGG分析表明circ_014154参与成骨细胞分化的正调控[54]。在小鼠胚胎成骨细胞前体细胞研究中发现,circ AFF4能够促进股骨骨折愈合[55]。此外circ FGFR2被证实能促进大鼠牙囊细胞成骨分化[10]。
3 、Circ RNAs在成骨分化调控中的信号通路研究
Wnt/β-catenin信号通路、丝裂原活化蛋白激酶(mitogen-activated protein kinases,MAPKs)通路、Notch信号通路和核因子-κB(NF-κB)信号通路等在组织成骨分化过程中发挥着不可替代的作用。随着对Circ RNAs研究的深入,Circ RNAs是否参与或影响重要信号通路的调控受到了广泛的关注。
3.1、 Wnt/β-catenin通路
Wnt/β-catenin信号通路(称为经典Wnt通路)广泛存在于多细胞动物中,是一条在物种进化中高度保守的信号通路,其构成主要包括Wnt、卷曲蛋白、β-catenin、低密度脂蛋白受体相关蛋白5/6、GSK-3β、T细胞因子/淋巴增强因子等[56]。目前,靶向Wnt/β-catenin信号通路已经成为维持骨内稳态、促进骨修复、治疗骨骼疏松的重要途径之一[57]。
已有大量研究证实,Circ RNAs可通过靶向广泛的mi RNA对Wnt/β-catenin信号通路的调控作用产生影响。据报道,沉默circ IGSF11可以促进BMSCs的成骨过程,并且与mi R-199b-5p呈负相关[37]。而mi R-199b-5p可以通过靶向糖原合成酶激酶3β(GSK-3β)刺激成骨细胞分化,其中GSK-3β可以通过诱导β-catenin进入细胞核,从而激活Wnt信号通路[58]。此外有报道表明,源自YAP1的circ_0024097使mi R-376b-3p上调了YAP1的水平,从而激活Wnt/β-catenin途径,进一步促进了BMSC和MC3T3-E1细胞的成骨分化[32]。
Qian等发现,circ RNA_5846与小鼠胚胎成骨细胞前体细胞成骨分化相关[12],且生物信息学分析表明circ RNA_5846与mi R-432之间存在相互作用。同时有研究表明,mi R-432可能通过下调Wnt/β-catenin信号通路来抑制人肝癌细胞的增殖[59]。但是否存在circ RNA_5846/mi R-432/Wnt/β-catenin途径调节成骨仍有待研究。
3.2 丝裂原活化蛋白激酶通路
MAPKs是普遍存在于各种哺乳动物细胞中的一组丝氨酸/苏氨酸蛋白激酶,能被多种外环境所激活,通过三级酶促级联反应进行信号转导,参与细胞生长、分化、凋亡等多种生理过程[60],尤其在BM-MSCs的成骨过程中起着重要作用。据报道,在HNGF6A处理后的小鼠胚胎成骨细胞前体细胞中,circ RNA_0001843可靶向结合mi R-214,进而抑制p38和JNK的磷酸化。此外,circ RNA_0001843过表达和mi R-214敲低均显着降低了HNGF6A的细胞保护和成骨促进作用。因此证明了HNGF6A通过靶向circ RNA_0001843/mi R-214通路及其下游激酶p38和JNK,保护成骨细胞免受氧化应激诱导的凋亡和成骨细胞表型抑制[53]。
此外报道,circ RNA_4214在成骨诱导的牙周膜干细胞中表达显着上调,并通过调控mi R-21及MAPKs信号通路促进成骨[51]。同时circ BANP、circ ITCH分别与mi R-34a和mi R-146a相互作用,通过MAPKs信号通路调节人牙周膜干细胞的成骨分化[48]。
3.3 、Notch信号通路
Notch信号抑制MSCs向成骨细胞系的分化,但在早期促进干细胞的增殖,在后期Notch激活促进干细胞成骨向分化[57]。有研究揭示,mi R-199b-5p过表达抑制C3H10T1/2细胞的生长,而促进了转化生长因子-β3诱导C3H10T1/2细胞向软骨细胞分化,增强软骨细胞特异性标志物SOX9、aggrecan和II型胶原(Col2a1)的基因和蛋白表达[61]。而沉默circ IGSF11被证实可以上调mi R-199B-5p的表达[37],因此推测circ IGSF11/mi R-199b-5p/JAG1轴可能具有调控C3H10T1/2细胞软骨向分化的潜能。
另外,Notch途径中重要的转录因子kappa J区重组信号结合蛋白(RBPJ)与DAB1启动子相互作用,而不与circ DAB1结合。circ DAB1过表达可导致RBPJ与DAB1启动子结合增强。circ DAB1通过结合和mi R-944上调RBPJ的表达水平。过表达circ DAB1可促进BM-MSCs的增殖和成骨分化。因此表明circ DAB1通过Noch/RBPJ途径促进BMSCs增殖和成骨分化[62]。
3.4 、其他信号通路
PI3K/AKT信号通路由Pl3K和下游效应器AKT构成,参与调控细胞存活、增殖、生长、分化等生理过程。有报道circ AFF4/mi R-7223-5p/PIK3R1轴在骨折愈合过程中具有促进作用[55]。
此外有学者发现circ RNA_3140与Toll样受体信号通路和NF-κB信号通路相关。circ RNA_3140可作为mi R-21的分子海绵,间接靶向激活素受体IIB在机械力诱导PDLSCs成骨分化中发挥关键作用[50]。
4 、Circ RNAs对成骨相关因子和蛋白的调控作用
4.1、 矮小相关基因RUNX
矮小相关转录因子,又称RUNX因子,蛋白质定位于亚核区域,并在基因启动子调控复合物的形成过程整合细胞信号[61]。哺乳动物的RUNX转录因子由RUNX1、RUNX2和RUNX3组成,其中RUNX2在胚胎发育和骨骼发育过程中起着至关重要的作用[63]。已有大量研究表明Circ RNAs能通过mi RNA海绵作用调控RUNX因子参与组织成骨分化过程,例如:circ RUNX2通过拮抗mi R-203,间接促进RUNX2的表达,从而发挥正向调控成骨分化的生物学功能[30]。在BMP2诱导的BM-MSCs成骨分化过程中circ RNA_33287作为mi R-214-3p的分子海绵,mi R-214-3p可靶向RUNX3的3’端非编码区来调控表达[11]。Xu等研究发现,circ_0011269在骨质疏松症患者标本中低表达,并验证了circ RNA_0011269/mi R-122和mi R-122/RUNX2的靶向关系,证实了circ RNA_0011269可以通过mi R-122调控RUNX2的表达水平[28]。同样,Han等发现circ_0076690可通过靶向mi R-152调控RUNX2,进而促进BMSCs的成骨分化[31]。
另外,在硅酸氢钙微粒诱导的BM-MSCs中敲低circ RNA_1983的表达水平后,mi R-6931表达水平上调,其靶基因Gas7和RUNX2的表达水平下降,成骨分化受到抑制[34]。circ RNA_0026827通过mi R-188-3p的分子海绵,间接调控下游BECLIN 1和RUNX1信号通路,进而促进人DPSCs向成骨细胞分化[43]。Yang等证实,人BM-MSCs中circ-VANGL1通过靶向mi RNA-217来调节RUNX2的表达,从而促进其成骨分化进程[27]。
4.2、 骨形态发生蛋白
骨形态发生蛋白(bone morphogenetic protein,BMP)信号通路在许多物种中高度保守,其在骨骼系统模式形成中具有重要性。BMP信号通路的紊乱会导致严重的骨缺损。这一途径是通过BMP配体与BMP受体I型和II型结合,进一步激活细胞内的Smads(Smad1、Smad5和Smad8)蛋白,Smads磷酸化可以与co-Smad4结合形成复合物,该复合物可以移位到细胞核并触发骨相关基因的表达[64]。
在降钙素基因相关肽诱导的小鼠胚胎成骨细胞前体细胞(MC3T3)中,mm9_circ_009056的表达上调,而mi R-22-3p的表达明显降低。沉默mm9_circ_009056可增加mi R-22-3p的表达,降低BMP7、RUNX2的基因和蛋白水平。后续实验进一步证实mm9_circ_009056可作为mi R-22-3p的分子海绵调控BMP7、RUNX2的表达,进而发挥促进小鼠胚胎成骨细胞前体细胞成骨分化的生物学作用[52]。此外,Ge等报道circ SIPA1L1通过吸附mi R-617影响下游靶基因Smad3的表达水平,进而促进DPSCs成骨[65]。
Chen等发现在大鼠牙囊细胞成骨分化过程中,circ FGFR2和BMP6表达增加,mi R-133表达降低。在此之前已有研究发现circ FGFR2可以通过用靶向结合海绵mi R-133a-5p和mi R-29b-1-5p促进骨骼肌增殖和分化[66]。因此猜想circ FGFR2/mi R-133/BMP6可能在牙齿再生和骨形成过程中发挥重要作用[10]。
4.3、 人源重组蛋白
人源重组蛋白(Nell-1)是一种分泌型成骨生长因子。circ RFWD2和circ INO80可以调节Hsa-mi R-6817-5p的表达,并影响重组NELL-1诱导的人脂肪干细胞的成骨分化[47]。骨不连患者的BM-MSCs中circ RNA_0074834的表达降低。circ RNA_0074834可作为ce RNA靶向mi R-942-5p调控ZEB1和VEGF的表达,进而促进BM-MSCs的成骨分化和骨缺损的修复[35]。
4.4 、整合素5
有报道称circ RNA-vgll3通过直接靶向mi R-326-5p调控ADSCs的成骨分化,mi R-326-5p通过抑制整合素5(integrin alpha5,ITGA5)的翻译而阻断成骨分化。ITGA5属于整合素受体家族,介导细胞粘附。在研究中,circ RNA-vgll3可以显着促进ITGA5的表达,因此,过表达circ RNA-vgll3的ADSCs可能具有更强的细胞粘附能力,包括归巢效应和骨祖细胞募集。此外,ITGA5与骨形成过程密切相关。在体内实验中,将ADSCs附载于CPC支架上,在过表达circ RNA-vgll3组中缺损区新骨形成水平显着增加,证实circ RNA-vgll3可增强骨的粘附、增加骨祖细胞募集和促进成骨分化[46]。
4.5 、骨糖蛋白
骨糖蛋白(Osteoglycin,OGN)也被称为骨诱导因子或Mimecan,是一种分泌到细胞外基质中的富含亮氨酸的小蛋白多糖,最初是从牛骨中分离出来的,作为基质矿化的诱导剂,在调节骨骼对改变能量平衡的适应性反应中起着关键作用[67]。Wen等发现,circ_0076906可以结合mi R-1305并调节其靶基因OGN的表达,从而调节人BM-MSCs的成骨分化,减缓骨质疏松的进展[26]。
4.6 、A激酶锚定蛋白2
A激酶锚定蛋白2(A-kinase anchoring protein2,AKAP2)可能参与信号通路中的极性形成或参与构建PKA-RII效应物复合物,以捕获、扩增和聚焦弥散性跨细胞c AMP信号传导系统[68]。许多研究揭示AKAP2的突变与骨疾病有关,其被鉴定为中国特发性脊柱侧弯家庭突变的新基因[68]。Lin等发现,抑制circ-SLC8A1可降低BMSCs矿化结节的形成,其机制主要是circ-SLC8A1可以充当ce RNA,通过mi RNA海绵作用调节mi R-516b-5p进一步调节AKAP2的表达,进而促进BMSCs的成骨分化[36]。
4.7、 Fo XO1因子
Fo XO1是FOXO的O类成员,是骨骼中酶抗氧化防御的主要调节剂,在成骨细胞中敲除Fo XO1会降低骨量和成骨细胞,而在成骨细胞中Fo XO3或Fo XO4不会影响骨量,研究表明Fo XO1是预防和治疗骨质疏松症的潜在靶标,Fo XO1通过调节成骨细胞的增殖和氧化还原平衡来促进骨形成[69]。在糖皮质激素诱发的骨质疏松症研究中发现,circ_0006393通过调控mi R-145-5p并上调Fo XO1来提高骨重塑过程中成骨基因的表达水平,进而促进细胞成骨分化,Gio P小鼠模型也证实了circ_0006393的促成骨潜能[33]。
5 、结语
目前骨缺损和骨质疏松等骨疾病的诊断及治疗方法仍在不断的更新迭代中,近年来Circ RNAs在生物矿化中所发挥的生物学作用已受到广泛研究及关注,但在基因表达及调控方面尚待解决的问题还有很多,例如在不同细胞系之间Circ RNAs-mi RNAs调控网络是否成立;在Circ RNAs调控网络中,各通路之间是否存在一定的协同作用;Circ RNAs-mi RNAs-m RNAs轴在成骨过程中调控方式以及其相互作用的具体分子机制。与此同时,Circ RNAs能否通过翻译蛋白或直接结合蛋白来调控成骨还有待研究。对Circ RNAs在成骨分化中的进一步深入研究将为骨缺损和骨质疏松等骨疾病的诊断和治疗提供新的思路。
参考文献
[1] Cocquerelle C,Mascrez B,Hétuin D,et al.Mis-splicing yields circular RNA molecules[J].FASEB J,1993,7(1):155-160.DOI:10.1096/fasebj.7.1.7678559.
[2] Sanger HL,Klotz G,Riesner D,et al.Viroids are singlestranded covalently closed circular RNA molecules existing as highly base-paired rod-like structures[J].Proc Natl Acad Sci USA,1976,73(11):3852-3856.DOI:10.1073/pnas.73.11.3852.
[3] Jeck WR,Sorrentino JA,Wang K,et al.Circular RNAs are abundant,conserved,and associated with ALUrepeats[J].RNA,2013,19(2):141-157.DOI:10.1261/rna.035667.112.
[4] Gla?ar P,Papavasileiou P,Rajewsky N.Circ Base:a database for circular RNAs[J].RNA,2014,20(11):1666-1670.DOI:10.1261/rna.043687.113.
[5] Rybak-Wolf A,Stottmeister C,Gla?ar P,et al.Circular RNAs in the mammalian brain are highly abundant,conserved,and dynamically expressed[J].Mol Cell,2015,58(5):870-885.DOI:10.1016/j.molcel.2015.03.027.
[6] Yu CY,Li TC,Wu YY,et al.The circular RNA circ BIRC6participates in the molecular circuitry controlling human pluripotency[J].Nat Commun,2017,8(1):1149.DOI:10.1038/s41467-017-01216-w.
[7] You X,Vlatkovic I,Babic A,et al.Neural circular RNAs are derived from synaptic genes and regulated by development and plasticity[J].Nat Neurosci,2015,18(4):603-610.DOI:10.1038/nn.3975.
[8] Han D,Li J,Wang H,et al.Circular RNA circ MTO1 acts as the sponge of micro RNA-9 to suppress hepatocellular carcinoma progression[J].Hepatology,2017,66(4):1151-1164.DOI:10.1002/hep.29270.
[9] Kristensen LS,Okholm TLH,Ven?MT,et al.Circular RNAs are abundantly expressed and upregulated during human epidermal stem cell differentiation[J].RNA Biol,2018,15(2):280-291.DOI:10.1080/15476286.2017.1409931.
[10] Du Y,Li J,Hou Y,et al.Alteration of circular RNAexpression in rat dental follicle cells during osteogenic differentiation[J].J Cell Biochem,2019,120(8):13289-13301.DOI:10.1002/jcb.28603.
[11] Peng W,Zhu S,Chen J,et al.Hsa_circ RNA_33287promotes the osteogenic differentiation of maxillary sinus membrane stem cells via mi R-214-3p/Runx3[J].Biomed Pharmacother,2019,109:1709-1717.DOI:10.1016/j.biopha.2018.10.159.
[12] Qian DY,Yan GB,Bai B,et al.Differential circ RNAexpression profiles during the BMP2-induced osteogenic differentiation of MC3T3-E1 cells[J].Biomed Pharmacother,2017,90:492-499.DOI:10.1016/j.biopha.2017.03.051.
[13] Zheng Y,Li X,Huang Y,et al.The circular RNAlandscape of periodontal ligament stem cells during osteogenesis[J].J Periodontol,2017,88(9):906-914.DOI:10.1902/jop.2017.170078.
[14] Meng S,Zhou H,Feng Z,et al.Circ RNA:functions and properties of a novel potential biomarker for cancer[J].Mol Cancer,2017,16(1):94.DOI:10.1186/s12943-017-0663-2.
[15] Li X,Yang L,Chen LL.The biogenesis,functions,and challenges of circular RNAs[J].Mol Cell,2018,71(3):428-442.DOI:10.1016/j.molcel.2018.06.034.
[16] Wesselhoeft RA,Kowalski PS,Anderson DG.Engineering circular RNA for potent and stable translation in eukaryotic cells[J].Nat Commun,2018,9(1):2629.DOI:10.1038/s41467-018-05096-6.
[17] Liang D,Wilusz JE.Short intronic repeat sequences facilitate circular RNA production[J].Genes Dev,2014,28(20):2233-2247.DOI:10.1101/gad.251926.114.
[18] D ong R,M a X K,Che n LL,et al.Increased complexity of circ RNA expression during species evolution[J].RNA Biol,2017,14(8):1064-1074.DOI:10.1080/15476286.2016.1269999.
[19] Werfel S,Nothjunge S,Schwarzmayr T,et al.Characterization of circular RNAs in human,mouse and rat hearts[J].J Mol Cell Cardiol,2016,98:103-107.DOI:10.1016/j.yjmcc.2016.07.007.
[20] Chen W,Schuman E.Circular RNAs in brain and other tissues:a functional enigma[J].Trends Neurosci,2016,39(9):597-604.DOI:10.1016/j.tins.2016.06.006.
[21] Bianco P,Cao X,Frenette PS,et al.The meaning,the sense and the significance:translating the science of mesenchymal stem cells into medicine[J].Nat Med,2013,19(1):35-42.DOI:10.1038/nm.3028.
[22] Yousefi AM,James PF,Akbarzadeh R,et al.Prospect of stem cells in bone tissue engineering:a review[J].Stem Cells Int,2016,2016:6180487.DOI:10.1155/2016/6180487.
[23] Zhang W,Dong R,Diao S,et al.Differential long noncoding RNA/m RNA expression profiling and functional network analysis during osteogenic differentiation of human bone marrow mesenchymal stem cells[J].Stem Cell Res Ther,2017,8(1):30.DOI:10.1186/s13287-017-0485-6.
[24] Wu Y,Lian K,Sun C.Lnc RNA LEF1-AS1 promotes osteogenic differentiation of dental pulp stem cells via sponging mi R-24-3p[J].Mol Cell Biochem,2020,475(1-2):161-169.DOI:10.1007/s11010-020-03868-7.
[25] Li M,Cong R,Yang L,et al.A novel lnc RNA LNC_000052leads to the dysfunction of osteoporotic BMSCs via the mi R-96-5p-PIK3R1 axis[J].Cell Death Dis,2020,11(9):795.DOI:10.1038/s41419-020-03006-7.
[26] Wen J,Guan Z,Yu B,et al.Circular RNA hsa_circ_0076906 competes with OGN for mi R-1305 biding site to alleviate the progression of osteoporosis[J].Int JBiochem Cell Biol,2020,122:105719.DOI:10.1016/j.biocel.2020.105719.
[27] Yang L,Zeng Z,Kang N,et al.Circ-VANGL1 promotes the progression of osteoporosis by absorbing mi RNA-217to regulate RUNX2 expression[J].Eur Rev Med Pharmacol Sci,2019,23(3):949-957.DOI:10.26355/eurrev_201902_16981.
[28] Xu X,Chen Y,Tan B,et al.Circular RNA circ_0011269sponges mi R-122 to regulate RUNX2 expression and promotes osteoporosis progression[J].J Cell Biochem,2020,121(12):4819-4826.DOI:10.1002/jcb.29709.
[29] Yu L,Liu Y.Circ RNA_0016624 could sponge mi R-98 to regulate BMP2 expression in postmenopausal osteoporosis[J].Biochem Biophys Res Commun,2019,516(2):546-550.DOI:10.1016/j.bbrc.2019.06.087
[30] Yin Q,Wang J,Fu Q,et al.Circ RUNX2 through has‐mi R‐203 regulates RUNX2 to prevent osteoporosis[J].JCell Mol Med,2018,22(12):6112-6121.DOI:10.1111/jcmm.13888.
[31] Han S,Kuang M,Sun C,et al.Circular RNA hsa_circ_0076690 acts as a prognostic biomarker in osteoporosis and regulates osteogenic differentiation of h BMSCs via sponging mi R-152[J].Aging (Albany NY),2020,12(14):15011-15020.DOI:10.18632/aging.103560.
[32] Huang Y,Xiao D,Huang S,et al.Circular RNA YAP1attenuates osteoporosis through up-regulation of YAP1and activation of Wnt/beta-catenin pathway[J].Biomed Pharmacother,2020,129:110365.DOI:10.1016/j.biopha.2020.110365.
[33] Wang XB,Li PB,Guo SF,et al.Circ RNA_0006393promotes osteogenesis in glucocorticoidinduced osteoporosis by sponging mi R1455p and upregulating FOXO1[J].Mol Med Rep,2019,20(3):2851-2858.DOI:10.3892/mmr.2019.10497.
[34] Zhong W,Li X,Pathak JL,et al.Dicalcium silicate microparticles modulate the differential expression of circ RNAs and m RNAs in BMSCs and promote osteogenesis via circ_1983-mi R-6931-Gas7 interaction[J].Biomater Sci,2020,8(13):3664-3677.DOI:10.1039/d0bm00459f.
[35] Ouyang Z,Tan T,Zhang X,et al.Circ RNA hsa_circ_0074834 promotes the osteogenesis-angiogenesis coupling process in bone mesenchymal stem cells (BMSCs)by acting as a ce RNA for mi R-942-5p[J].Cell Death Dis,2019,10(12):932.DOI:10.1038/s41419-019-2161-5.
[36] Lin C,Zhong W,Yan W,et al.Circ-SLC8A1 regulates osteoporosis through blocking the inhibitory effect of mi R-516b-5p on AKAP2 expression[J].J Gene Med,2020,22(11):e3263.DOI:10.1002/jgm.3263.
[37] Zhang M,Jia L,Zheng Y.Circ RNA expression profiles in human bone marrow stem cells undergoing osteoblast differentiation[J].Stem Cell Rev Rep,2019,15(1):126-138.DOI:10.1007/s12015-018-9841-x.
[38] Kuang MJ,Xing F,Wang D,et al.Circ USP45 inhibited osteogenesis in glucocorticoid-induced osteonecrosis of femoral head by sponging mi R-127-5p through PTEN/AKT signal pathway:experimental studies[J].Biochem Biophys Res Commun,2019.509(1):255-261.DOI:10.1016/j.bbrc.2018.12.116.
[39] Zhu S,Zhu Y,Wang Z,et al.Bioinformatics analysis and identification of circular RNAs promoting the osteogenic differentiation of human bone marrow mesenchymal stem cells on titanium treated by surface mechanical attrition[J].Peer J,2020,8:e9292.DOI:10.7717/peerj.9292.
[40] Ge X,Li Z,Jing S,et al.Parathyroid hormone enhances the osteo/odontogenic differentiation of dental pulp stem cells via ERK and P38 MAPK pathways[J].J Cell Physiol,2020,235(2):1209-1221.DOI:10.1002/jcp.29034.
[41] Li C,Jiang H.Altered expression of circular RNAin human dental pulp cells during odontogenic differentiation[J].Mol Med Rep,2019,20(2):871-878.DOI:10.3892/mmr.2019.10359.
[42] Ferrarotti F,Romano F,Gamba MN,et al.Human intrabony defect regeneration with micrografts containing dental pulp stem cells:a randomized controlled clinical trial[J].J Clin Periodontol,2018,45(7):841-850.DOI:10.1111/jcpe.12931.
[43] Ji F,Zhu L,Pan J,et al.Hsa_circ_0026827 promotes osteoblast differentiation of human dental pulp stem cells through the beclin1 and RUNX1 signaling pathways by sponging mi R-188-3p[J].Front Cell Dev Biol,2020,8:470.DOI:10.3389/fcell.2020.00470.
[44] Laloze J,Fiévet L,Desmoulière A.Adipose-derived stromal cell in regenerative medicine:a review[J].Adv Wound Care (New Rochelle),2021,10(1):24-28.DOI:10.1089/wound.2020.1175.
[45] Betz VM,Kochanek S,Rammelt S,et al.Recent advances in gene‐enhanced bone tissue engineering[J].The Journal of Gene Medicine,2018,20(6):e3018.DOI:10.1002/jgm.3018.
[46] Zhang D,Ni N,Wang Y,et al.Circ RNA-vgll3 promotes osteogenic differentiation of adipose-derived mesenchymal stem cells via modulating mi RNA-dependent integrinα5expression[J].Cell Death Differ,2021,28(1):283-302.DOI:10.1038/s41418-020-0600-6.
[47] Huang X,Cen X,Zhang B,et al.The roles of circ RFWD2and circ INO80 during NELL‐1‐induced osteogenesis[J].J Cell Mol Med,2019,23(12):8432-8441.DOI:10.1111/jcmm.14726.
[48] Gu X,Li M,Jin Y,et al.Identification and integrated analysis of differentially expressed lnc RNAs and circ RNAs reveal the potential ce RNA networks during PDLSCosteogenic differentiation[J].BMC Genet,2017,18(1):100.DOI:10.1186/s12863-017-0569-4.
[49] Li X,Zheng Y,Zheng Y,et al.Circular RNA CDR1as regulates osteoblastic differentiation of periodontal ligament stem cells via the mi R-7/GDF5/SMAD and p38MAPK signaling pathway[J].Stem Cell Res Ther,2018,9(1):232.DOI:10.1186/s13287-018-0976-0.
[50] Wang H,Feng C,Jin Y,et al.Identification and characterization of circular RNAs involved in mechanical force‐induced periodontal ligament stem cells[J].J Cell Physiol,2019,234(7):10166-10177.DOI:10.1002/jcp.27686.
[51] Quarles LD,Yohay DA,Lever LW,et al.Distinct proliferative and differentiated stages of murine MC3T3‐E1 cells in culture:an in vitro model of osteoblast development[J].J Bone Miner Res,1992,7(6):683-692.DOI:10.1002/jbmr.5650070613.
[52] Wu C,Zheng Z,Ren W,et al.Mm9_circ_009056 enhances osteogenesis by targeting BMP7 via CGRP-mediated mi R-22-3p[J].Biochem Biophys Res Commun,2018,501(1):199-205.DOI:10.1016/j.bbrc.2018.04.215.
[53] Zhu X,Zhao Z,Zeng C,et al.HNGF6A inhibits oxidative stress-induced MC3T3-E1 cell apoptosis and osteoblast phenotype inhibition by targeting circ_0001843/mir-214pathway[J].Calcif Tissue Int,2020,106(5):518-532.DOI:10.1007/s00223-020-00660-z.
[54] Cao Z,Zhang Y,Wei S,et al.Comprehensive circ RNAexpression profile and function network in osteoblast-like cells under simulated microgravity[J].Gene,2021,764:145106.DOI:10.1016/j.gene.2020.145106.
[55] Mi B,Xiong Y,Chen L,et al.Circ RNA AFF4 promotes osteoblast cells proliferation and inhibits apoptosis via the Mir-7223-5p/PIK3R1 axis[J].Aging (Albany NY),2019,11(24):11988-12001.DOI:10.18632/aging.102524.
[56] Huang P,Yan R,Zhang X,et al.Activating Wnt/β-catenin signaling pathway for disease therapy:challenges and opportunities[J].Pharmacol Ther,2019,196:79-90.DOI:10.1016/j.pharmthera.2018.11.008.
[57] Baron R,Kneissel M.WNT signaling in bone homeostasis and disease:from human mutations to treatments[J].Nat Med,2013,19(2):179-192.DOI:10.1038/nm.3074.
[58] Zhao R,Li Y,Lin Z,et al.mi R-199b-5p modulates BMSC osteogenesis via suppressing GSK-3β/β-catenin signaling pathway[J].Biochem Biophys Res Commun,2016,477(4):749-754.DOI:10.1016/j.bbrc.2016.06.130.
[59] Jiang N,Chen WJ,Zhang JW,et al.Downregulation of mi R-432 activates Wnt/β-catenin signaling and promotes human hepatocellular carcinoma proliferation[J].Oncotarget,2015,6(10):7866-7879.DOI:10.18632/oncotarget.3492.
[60] Kim HY,Park SY,Choung SY.Enhancing effects of myricetin on the osteogenic differentiation of human periodontal ligament stem cells via BMP-2/Smad and ERK/JNK/p38 mitogen-activated protein kinase signaling pathway[J].Eur J Pharmacol,2018,834:84-91.DOI:10.1016/j.ejphar.2018.07.012.
[61] Zhang M,Yuan SZ,Sun H,et al.mi R-199b-5p promoted chondrogenic differentiation of C3H10T1/2 cells by regulating JAG1[J].J Tissue Eng Regen Med,2020,14(11):1618-1629.DOI:10.1002/term.3122.
[62] Chia W,Liu J,Huang YG,et al.A circular RNA derived from DAB1 promotes cell proliferation and osteogenic differentiation of BMSCs via RBPJ/DAB1 axis[J].Cell Death Dis,2020,11(5):372.DOI:10.1038/s41419-020-2572-3.
[63] Mevel R,Draper JE,Lie-A-Ling M,et al.RUNXtranscription factors:orchestrators of development[J].Development,2019,146(17):dev148296.DOI:10.1242/dev.148296.
[64] Chen G,Xu H,Yao Y,et al.BMP signaling in the development and regeneration of cranium bones and maintenance of calvarial stem cells[J].Front Cell Dev Biol,2020,8:135.DOI:10.3389/fcell.2020.00135.
[65] Ge X,Li Z,Zhou Z,et al.Circular RNA SIPA1L1 promotes osteogenesis via regulating the mi R-617/Smad3 axis in dental pulp stem cells[J].Stem Cell Res Ther,2020,11(1):364.DOI:10.1186/s13287-020-01877-3.
[66] Chen X,Ouyang H,Wang Z,et al.A novel circular RNAgenerated by FGFR2 gene promotes myoblast proliferation and differentiation by sponging mi R-133a-5p and mi R-29b-1-5p[J].Cells,2018,7(11):199.DOI:10.3390/cells7110199.
[67] Lee NJ,Ali N,Zhang L,et al.Osteoglycin,a novel coordinator of bone and glucose homeostasis[J].Mol Metab,2018,13:30-44.DOI:10.1016/j.molmet.2018.05.004.
[68] Li W,Li Y,Zhang L,et al.AKAP2 identified as a novel gene mutated in a Chinese family with adolescent idiopathic scoliosis[J].J Med Genet,2016,53(7):488-493.DOI:10.1136/jmedgenet-2015-103684.
[69] Liao L,Su X,Yang X,et al.TNF-αinhibits Fox O1 by upregulating mi R-705 to aggravate oxidative damage in bone marrow-derived mesenchymal stem cells during osteoporosis[J].Stem Cells,2016,34(4):1054-1067.DOI:10.1002/stem.2274.





