
���������ͷ������������е�����ʧ��DZ��;��. ���������ļӾ�, �����е������ͷ��������û���������ͷŸ�����������������ǵ�. ����Ϊ��������������ǵ���, �����������ͷ���������65%����[1]. ����������(denitrification)��ָ�������εȽϸ��ӵĺ���������ת��Ϊ���� �� һ�������Ͷ��������Ĺ���, ��������������(>60%��������϶ˮ����)½�������ǵ���������Ҫ�������. �ڹ�ȥ�� 1 ����������, ����ͨ����Ϊ���������̶����ɽ�������������ϸ�����е�, �����������о�����, ���Ҳ���Խ��з���������[2]. �ڷ����õ������, �Ѿ����ֺܶ��־��в��������ǵ�������.
������Ȼ�ڴ�����������, ������������ǵ�������ϸ�����Ҫ�ͼ���������[3], ���Ƕ����������ǵ����������IJⶨ�������, ����������ǵ�������Զ��ϸ��[4-5]. Laughlin ��[4]�о�����: �ڲݵؼ�������̿����ʹ�÷��߾�ͪ�����������ʱ, ���������ǵ��ŷ�ͨ�������� 80%, ������ù����������ϸ�����Ժ�, ���������ǵ��ŷ�ͨ�������� 30%����. ��������������ǵ��ŷ��е���Ҫ��������������������������������ռ���ŷdz��������. ����ԭλ�����ǵ��ŷ�ͨ����������������������������,���뷴����ϸ����������������û�����Ե����[6]. �Ѿ���һЩ���ڲݵء� ʪ���Լ����ص���̬ϵͳ������������ǵ��ŷŵ��о�, ���Ƕ����ֵ��������о��в����[4-5, 7-8]. ���־����ֵļ�Լ��Ӫ�̶�Խ��Խ��, ���Ƕ���������, �ر��Ƕ���������ĸ���Ҳ�dz�����. ��������������ܸ��͵� pH ֵ,�����ֵ������е�������Ҳ��������ϸ��, ���������Ϸ�����ɭ�����������ǵ��ŷŹ����п��ܾ��и���Ĺ���[9-10]. ë�� Phyllostachy edulis ���й��Ϸ�����Ҫ����[11], ���¸�����, ��������ǿ, ������ë�����ܱ�ɭ�����Ӳ����ԭ��ɭ��. ����, ë���ֵľ��ü�ֵ�ϸ�, Ŀǰũ������ļ�Լ��Ӫ�̶�Ҳ�ڲ������. �����������Ϊ���Ž�Ϊ����, ԭ����Ȼ�ָı�Ϊë���ֱؽ������������Ⱥ��ṹ�ı仯, �����������������ǵ��ŷ�ͨ���ĸı�. ���о�����ѡ�������Ʒ���, �о�ë���ּ���Ҷ������ϸ��������������ǵ��ŷŵĹ�����, ��ʾ��ͬ�������ֵ��������������ŷ��е����ü���Ӱ�����, �Ӷ�����Ҷ���ݱ��ë���ֽ�����̬��������, Ϊ������Ӫë����, �������������ŷ��ṩ��������.
����1 �����뷽��
����1.1 ��Ʒ�ɼ�
�������㽭ʡ�ٰ�����Ŀɽ���Ҽ���Ȼ����������Ұ�����, �����Ƶ��ε�ò�����·ֱ�ѡȡ 4 ��ë���ֵ�(moso bamboo forest, MB)�� 4 ����Ҷ�ֵ�(broad-leaved forest, BL). ��������������Ϊ�����ҷ����ĺ���, ë���ֵ�����Ϊ�����(��� 43.27%, ���� 40.34%, ɰ�� 16.39%), ��Ҷ�ֵ�����Ϊ�������(��� 38.50%, ���� 48.68%, ɰ�� 12.82%). ���� 5 ���������ȡ����, �ɼ�����(0~20 cm)����,װ���Է������ʵ����. ȥ����Ķ�ֲ����弰ʯ����, ��Ʒ�� 2 mm ��ɸ����, �ķַ�����ȥ������Ʒ. ��Ʒ��Ϊ 2 ��, 1 �������������������������֬֬�������, �� 1 �ݷ�ɺ�ⶨ���� pH ֵ�� �л��ʡ� ȫ�����������Ȼ�����������.
����1.2 ��������
������Ϻ��������������������������, �ⶨϸ������������������ǵ��ŷ�ͨ���Ĺ�����. ���鹲��Ϊ 4 ������: �ٶ���(control), �������κο�����; ����ù��(streptomycin, strep), ���������� 1.0mg·g-^1����ù��;�۷��߾�ͪ(cycloheximide, cyclo), �������� 2.0 mg·g-1���߾�ͪ;����ù��+���߾�ͪ(both), ������ͬʱ���� 1.0 mg·g^-1����ù�غ� 2.0 mg·g^-1���߾�ͪ. �ظ� 3 ��·����^-1.
����1.3 ���鷽��
����1.3.1 ������������ ��ȡ�൱�� 20.0 g �����������������Ž� 150 mL ��ɫ�Լ�ƿ��, ���� 4.0 mL ��ͬ��������Һ(���ս���ȥ����ˮ)ʹ�������������������ﵽ�������Ҫ��. �Լ�ƿװ��������, ����4 �������¹�ҹԤ����, ��ʹ����������ɢ������������϶��. ֮��, ���Լ�ƿ������������, �������зֱ���� 100 mmol·L^-1�����Ǻ� 100 mmol·L^-1�����, ʹ������ˮ���ﵽ�������ˮ���� 90%����. ���Լ�ƿ������������, �����г��������Ϊ 10%����Ȳ�����������ǵ�������ԭ�ɵ���. ����������Һ��ֻ�Ϻ�, ������������ 18 ������ 7 h ��, ��ʼ����Ͳ��ȡ�Լ�ƿ������, ����(GC-2014)����ɫ�ײⶨ�����ǵ�ͨ��.
����1.3.2 ��֬֬������� ������Ʒ����ʵ���Һ�, ���������䶳����, �ⶨ������֬֬����Ħ������Ũ��. ��֬֬����ⶨ���� Wu ��[12]����. ��ȡ 3.0 g ��������, �����ȷ���ȡ����֬��, ����Ʒװ�ع轺��(SPE-Si), �ֱ����ȷ¡� ��ͪ�ͼ״�ϴ������֬�� ��֬�ͼ�����֬. ��������ֽ��м���, �ȷ»���, �������ɺ�����ɫ��(������ 6890N)�ⶨ. ����֬֬����Ħ������Ũ�����ڱ���������������. ϸ���Ĵ��������Ϊ: i15:0, a15:0, 15:0, i16:0, 16:1w7c, i17:0, a17:0, cy17:0, 17:0, 18:1w7c, cy19:0w8c; ���������֬����Ϊ 18:2w6,9c[13]. ��֬֬����Ħ������Ũ���� nmol·g-1������ʾ.
����1.3.3 �����������ʷ��� �����л��ʲ����ظ��������ȷ��ⶨ; ȫ�����ÿ��Ϸ��ⶨ; ��̬�����̬�������Ȼ��ؽ���, ���������Dzⶨ; ��Ч�ײ��� Bray ���ⶨ, ����-�������Һ����, ��ɫ���ⶨ; ��Ч�ز����������ȡ-�����ȷ��ⶨ; pH ֵ���� 1��2.5 ��ˮ��, ��ȼƲⶨ[14].
����1.4 ���ݷ���
�������� Excel 2007 �� SPSS 18.0 ��������, Duncan �����ط���������Ƚϸ�����֮��IJ���������(P <0.05), ������� t ����Ƚ�ë��������Ҷ�������������ʼ�����ϸ������������������IJ���������, ���� Pearson ���ϵ�����鲻ͬ���������ǵ��ŷ������������������ʵ���ع�ϵ.
����2 ��������
����2.1 ������������
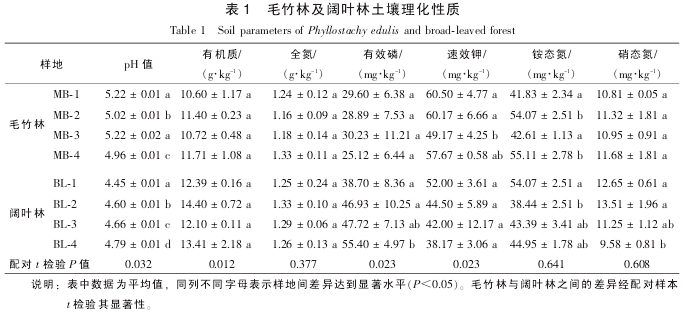
���������������������ֳ���ͬ�̶ȵ�����, ����ͬë���ּ���Ҷ������֮������ pH ֵ����Ҳ�ϴ�.
���������������: ë���ֲ�ͬ���������л��ʡ� ȫ���� ��Ч����̬������������û�����Ų���, ����Ҷ�ֲ�ͬ����֮�������л��ʡ� ȫ������Ч��Ҳû�����Ų���, ˵����ͬ�ַ������л��ʼ�ȫ�����������ֲ������Ϊ��һ. �Ա� 2 ���ַ�, ��Ҷ������ pH ֵ���ŵ���ë����(P<0.05), ���л�������Ч�������������Ÿ���ë��������(P<0.05). �෴, ë����������Ч�������������Ÿ�����Ҷ��(P<0.05,�� 1). ë����������Ҷ�ֻ���Ҷ���˹�����Ϊë���ֺ�, ���ڵ���ֲ���Լ�����������ĸı�, ������������Ҳ��������Ӧ�ı仯. ����, ë������Ϊ���ų̶�Ҳ�ڲ�������, �����ࡢ ������ ʩ���Լ������, ���ᵼ�����ֺ����ĸı�, �������л�����������. Ф������[15]�о�����: ���ڼ�Լ��Ӫ��, ë���������л�̼���½�. ��ë����������Ч�ظ�����Ҷ��, ������������Ϊʩ������ɵ�.
����2.2 ����������
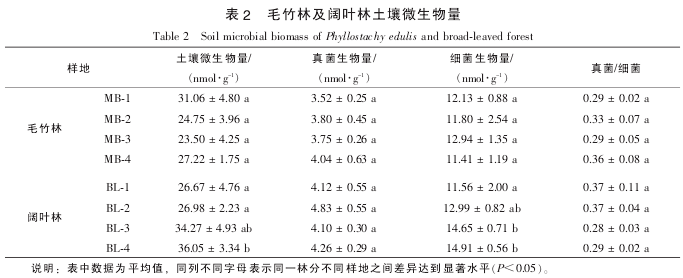
������֬֬�����Ǽ������л�������ϸ��Ĥ����Ҫ�ɷ�, ��ת���ʼ�������ϸ��������Ѹ�ٽ���[16].���, ���������������������֬֬����Ħ������Ũ����Ϊ������������ָ��, ��������ȵزⶨ�����еĻ�������. �������: ���������������� ϸ��������������Լ����/ϸ����ֵ, ë��������Ҷ�� 2 ���ַ�֮���û�����Ų���. ͬ��, ë���ֵIJ�ͬ����֮��Ҳû�����Ų���. ��Ҷ��������, BL-4 ��������������ϸ�������������Ÿ��� BL^-1 ����(P<0.05, �� 2). Pearson ��ط�������:
������������������� pH ֵ�����Ÿ����(R²=-0.752, P<0.05), ���������л��ʳʼ���������� (R²=0.965, P<0.01); ϸ����������������Ч���������(R²=0.830, P<0.05), ���������/ϸ����ֵ����̬���������(R²=0.815, P<0.05). ��ϸ�����, ���������Ӧ����� pH ֵ��Χ, ������ pH ֵ�����������������ϸ��������[17]. ���, �����Ե�ë���ֺ���Ҷ��������, ����Ļ�����Խ�ǿ. �л���Ҳ��Ӱ�������һ����Ҫ����. �о�����: ɭ��ת��Ϊ����֮���������л��ʽ���, ���������ֵ��������������[18]. �������л������Ƶ���, ë�����������������Ҷ��������½�������, ����û�����Ų��졣
����2.3 ϸ������������ǵ��ŷŹ���
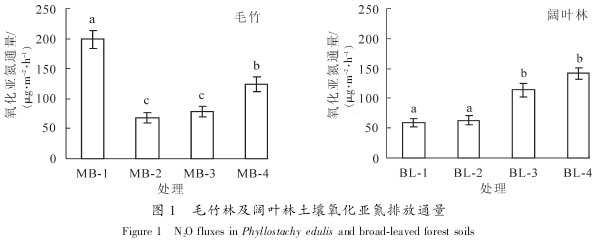
������ͬ�ַ������Լ�ͬһ���ֲַ�ͬ����֮�������ǵ����ŷ�ͨ�������ڽϴ����, ����ë���� MB-1 �������������ǵ��ŷ�ͨ�����, ����� MB-4, BL-3 �� BL-4 ������, MB-2 �� BL^-1, BL-2 �����ص������ǵ��ŷ�ͨ�����(ͼ 1). ��ط����������: ���������ǵ��ŷ�ͨ���뱾�о���ѡ��ⶨ�IJ���������������û�����������. ���������ǵ��ŷ������������й�, ��������ˮ���� ���ء� pH ֵ�� �л������������� ����ʩ�����ȵ�[19]. ���, ���о��в�ͬ�ַ�����ͬһ�ֲַ�ͬ����֮����ڵľ����, �������ɶ��������������ع�ͬ������, ��Ӱ�����ػ��д��ڽ�һ�����о�. ͳ�ƽ������: ë���ֺ���Ҷ�����������ǵ��ŷ�ͨ��û�����Ų���(t=-0.574, P=0.606). ��������ɭ�����������ǵ��ŷ�ͨ�����о�������, �����������¶Ⱥ�ʪ����Ϊ�ؼ�����[20], �����������о�����: ����ˮƽ��һ���ؼ�����[21]. �Ա� 2 �ֲ�ͬ�ַ�, ����������ȫ�����ǿ���̬����û�����Ų���(�� 1), ������� 2 ���ַ������ǵ��ŷ�ͨ��û�в������Ҫԭ��.
���������ǵ������ӷ��߾�ͪ�� ��ù�ػ���ͬʱ���� 2 �ֿ�����, ���������������������ǵ��ŷ�ͨ��(P<0.05). ͬʱ���� 2 �ֿ����ض����������ǵ��ŷ�ͨ������������ 73%~83%, ����ͬһ�ֵַIJ�ͬ����֮��û�����Ų���, ������������ù�ػ��߷��߾�ͪ�Ĵ��������������ǵ��ŷ�ͨ���������ʱ仯�ϴ�. ͼ 2 �������: �� 2 ���ֵַ�������, ���˸���������(MB-2, BL^-1), ��������������������ǵ��ŷ�ͨ�������Ÿ���ϸ��(P<0.05). ����ë����������, ��������������ǵ��ŷŵĹ�����Ϊ40.8%~56.2%, Զ����ϸ��(9.0%~33.4%). ��Ҷ��������Ҳ����ͬ���Ĺ���, ��������������ǵ��ŷŹ�����Ϊ 38.0%~62.8%, ��ϸ���Ĺ�����Ϊ 12.6%~32.4%(ͼ 2). Ŀǰ, �����о�����, ��ɭ����̬ϵͳ�е����Ⱥ������������ǵ��ŷŵĹ���Ҫ���ڲݵ��Լ���Ϊ��������̬ϵͳ[4-5, 8]. ͬʱ, �����ϸ�������������ǵ��ŷŵĹ��ױ��ڲ�ͬ����̬ϵͳ�д����Žϴ�IJ���[22]. ë��������Ҷ�����������ֲ���Լ���Ϊ���ų̶Ȳ�һ��, �����ϸ�������������ǵ��ŷŵĹ�����Ҳ�ʹ�����һ���IJ���.
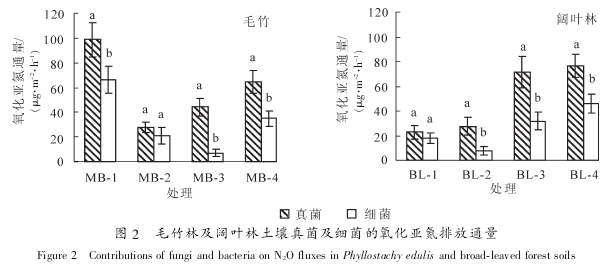
����������Ƶ���, �� 2 ���ֵַIJ�ͬ����֮��, ���������ϸ���������ǵ��ŷ�ͨ���Ĺ�����Ҳ�������ŵIJ���(P<0.05). ����ë����������, ���������ǵ��ŷ�ͨ�����ŵ������� MB-1(P<0.05), ����MB-3 ���ص�������������ǵ��ŷŹ����ʴﵽ 56.2%, ���Ÿ��� MB-1 ��������ŷŹ�����(49.7%,P<0.05). ������Ҷ��������, BL-3 �� BL-4 ���������������ǵ��ŷ�ͨ�����Ÿ���(P<0.05)���� 2 �����ص�ͬʱ, ����������ǵ��ŷŹ�����Ҳ���Ÿ������� BL^-1 �� BL-2(P<0.05).�����������ǵ��ŷ�ͨ�������������������ʼ��������������� Pearson ��ط���. �������:
���������ϸ���������ǵ��ŷŹ���֮����ڼ����ŵ������(P<0.01), ������ 2 ������Ⱥ�䷴�����Ļ�������Ҫ���Ϊ�����й�. ֵ��ע�����, �����ϸ�������ǵ��ŷ�ͨ������������֮��û�����������, ˵�������������Ǿ��������ǵ��ŷ�ͨ���Ĺؼ�����. ����ֻ��һЩ�ض��������ϸ��������Խ��з���������, ������е��������� Fusarium spp.���н�ǿ�������ǵ��ͷ�����[23]. ���, ����������ǵ��ŷŹ�����Ҫ�����Ⱥ�估�ض���Ⱥ�ķ������������. �����Ȼ��Ҫ���������ֶ������Լ����ܻ����ȵȷ�������һ���о��������������õ���Ҫ��������ؼ�Ӱ������. �������о��������: �����ϸ���������ǵ��������������� pH ֵ��ϵ�ϴ�, ����������� pH ֵ���� 2 ���������������Ծ�������Ҫ��Ӱ��[22, 24]. �������������Ӧ�dz����� pH ֵ��Χ, �������� pH ֵ�ı��Ӱ�����ϸ��С, ��ϸ�������������������¿��ܻ��ܵ�����, �������������������������ǵ��ŷŵĹ��ͻ����ϸ��. Ȼ�������������ͬ����, ���о��������������ϸ�������ǵ��ŷ�ͨ�������� pH ֵû��������ع�ϵ, ������벻ͬ����֮�� pH ֵ��Ϊ�ӽ��й�. ����, ��������ǵ��ŷ�ͨ����������̬�������н�ǿ�ĸ���ع�ϵ, ���ܲ�������(R²=-0.673, P=0.067). ͨ��, �����ǵ�ͨ���� NO3-N�ʸ���ع�ϵ. ���ŷ��������õĽ���, NO3-N ��Ϊ�������庬���½�, ��ԭ�������ǵ��ͷų���[25].
����3 ������չ��
����������ѡ������������������: ��������ë���ֻ�����Ҷ��, ����������������ǵ��ŷŵ���Ҫ������; ����Ҷ������Ϊë���ּ�û�иı����������ǵ��ŷ�ͨ��, Ҳû��Ӱ�������ϸ�������������ǵ��ŷ�ͨ������Թ�����; �������ϸ�������ǵ��ŷ���������, ����������û�������.
����������, ë���ֲ��������Ҷ�ֵ�ͬʱ, ��Ӫǿ��Ҳ�ڲ�������. ��ͳ��, �� 40%~50%��ë����ʵ�м�Լ������. ����Ȼ״̬�Ĵַž�Ӫë�������, ��Լ��Ӫë���ֵĵ����ص����Ϊ��������Ӳ�, ���������Լ�ʩ�ô������ʵ�. ���ܼ�Լ��Ӫ��ʩ����һ���̶�������ë����ϲ���������, ���̼����[26], ��Ҳ���о�����, ʹ�û�������������ë��������������̼�������ǵ����ŷ�ͨ��, ��ë������������������ŷ��������Ż���ʩ�������ӳ�������������[27], ���������ڻ����Բ����. ��Ϊ�����緭���� �����Լ�����ʩ�õȶ���Ӱ���������Ⱥ��ṹ����[28], �̶������������ǵ��ŷŲ���DZ��Ӱ��. ���о���, ����ֻ�������Ҷ�ֺ�ë���ֽ������о�, ���Dz�ͬ��Ӫ�����Լ���Ӫǿ�ȶ�ë����������������ǵ�����ŷŹ���Ӱ����δ�漰, ��������ڸ��ӹ㷺�ķ�Χ������˹���Ӫë���ֽ��в���, ������ȶ���ͬλ�ر���ֶ����о������ɭ��, �ر����˹���Ӫɭ����̬ϵͳ�����ǵ��ŷ��еĹ���. ��ȷ������������ǵ��ŷ���Թ������ë������Ϊ��Ӫ����Ӧ����, ���ں�����Ӫë����, ����ȫ������仯������ʮ����Ҫ������.
���������:
����[1] SMITH K A, CONEN F. Impacts of land management on fluxes of trace greenhouse gases [J]. Soil Use Manage,2004, 20(2): 255 - 263.
����[2] ZUMFT W G. Cell biology and molecular basis of denitrification [J]. Microbiol Mol Biol Rev, 1997, 61(4): 533 - 616.
����[3] KURAKOV A V, NOSIKOV A N, SKRYNNIKOVA E V, et al. Nitrate reductase and nitrous oxide production byFusarium oxysporum 11dn1 under aerobic and anaerobic conditions [J]. Curr Microbiol, 2000, 41(2): 114 - 119.
����[4] LAUGHLIN R J, STEVENS R J. Evidence for fungal dominance of denitrification and co-denitrification in agrassland soil [J]. Soil Sci Soc Am J, 2002, 66(5): 1540 - 1548.
����[5] CRENSHAW C L, LAUBER C, SINSABAUGH R L, et al. Fungal control of nitrous oxide production in semiaridgrassland [J]. Biogeochemistry, 2008, 87(1): 17 - 27.
����[6] KOMATSUZAKI M, SATO Y, OHTA H. Relationships between fungal biomass and nitrous oxide emission in uplandrice soils under no tillage and cover cropping systems [J]. Microbes Environ, 2008, 23(3): 201 - 208.
����[7] KINNEY C A, MOSIER A R, FERRER I, et al. Effects of the herbicides prosulfuron and metolachlor on fluxes ofCO2, N2O, and CH4in a fertilized Colorado grassland soil [J]. J Geophys Res, 2004, 10 9 (D5): doi: 10.1029/2003JD003536.
����[8] MCLAIN J E T, MARTENS D A. N2O production by heterotrophic N transformations in a semiarid soil [J]. Appl SoilEcol, 2006, 32(2): 253 - 263.
����[9] JOERGENSEN R G, WICHERN F. Quantitative assessment of the fungal contribution to microbial tissue in soil [J].Soil Biol Biochem, 2008, 40(12): 2977 - 2991.
����[10] STRICKLAND M S, ROUSK J. Considering fungal:bacterial dominance in soils-methods, controls, and ecosystemimplications [J]. Soil Biol Biochem, 2010, 42(9): 1385 - 1395.
����[11] ������, ���﷼, ������, ��. ��Ŀɽë��������Ҷ�ֺ�����ϸ��Ⱥ�� 16S rDNA V3 ��Ƭ�� PCR �� DGGE ����[J]. ����ѧ��, 2009, 46(4): 662 - 669.WANG Qizan, XU Qiufang, JIANG Peikun, et al. DGGE analysis of PCR of 16SrDNA V3 fragment of soil bacteriacommunity in soil under natural broadleaf forest invaded by Phyllostachy pubescens in Tianmu Mountain NatureReserve [J]. Acta Pedol Sin, 2009, 46(4): 662 - 669.





