
棉花粉蚧 Phenacoccus solenopsis Tinsley,又名扶桑绵粉蚧,2008 年在我国广东省广州市首次发现该虫入侵危害( 武三安和张润志,2009) 。该虫繁殖能力强、年发生世代多、寄主范围宽( 陆永跃等,2008;Joshi et al.,2010; Khuhro et al.,2011) 。在我国棉花粉蚧正快速向更广泛地区扩散传播,截至 2012 年12 月已在广东、海南、广西等 11 个省市区 100 余个县区发现该虫发生为害( 农业部农办农[2013]43号,2013) 。棉花粉蚧对棉花为害严重 ( Abbaset al.,2005; 孙峰和陆永跃,2011 ) ,对亚洲和我国的棉花产业潜在威胁严重( Wang et al.,2010) 。基于 CLIMEX 模型对棉花粉蚧在中国潜在地理分布预测发现,该虫潜在分布区域广泛,覆盖了除西藏、青海、黑龙江 3 个省区大部地区以外的所有其他区域,适宜分布区包括了长江中下游、黄河中下游、西北内陆 3 大主要棉区以及华南和辽河流域的零星棉区所有地区( 马骏等,2011) ,研究显示该虫过冷却点低,1 龄若虫达 - 24℃ ,耐寒性较强,如经低温锻炼后可能会更低,可能适宜在我国北部更广泛区域生存( 关鑫等,2009) 。
光周期对昆虫的生长发育与繁殖有重要作用( Roditakis et al.,2001; Keena et al.,2012; Kollberget al.,2013; Zerbino et al.,2013) 。光周期会影响植食性昆虫的取食表现( Beck,1980) 。伴随着环境因素的剧烈变化,光周期将成为影响物种调节生命周期的关键因素; 光周期常是决定昆虫滞育过程的关键因素之一,其诱导产生的滞育不仅是昆虫克服不良环境条件的一种适应机制,也是昆虫保持其生活史与季节同步的一种策略( 赵洪霞等,2011) 。光周期对昆虫种群适应性分化也有一定影响: 18℃、光周期 8L∶ 16D 和 10L∶ 14D 条件诱导高纬度棉蚜Aphis gossypii 种群产生了性蚜,而对低纬度种群无诱导效应( 刘健等,2003) ; 长光照条件下水稻叶蝉Trigonotylus caelestialium 雌成虫产生非滞育卵,短光照条件下产生滞育卵( Shintani,2009) 。
目前,关于环境因素影响棉花粉蚧种群的研究主要集中在温度( Patil et al.,2011; Lu et al.,2011;王莹莹等,2012; Guan et al.,2012; Kedar et al.,2012; Ali et al.,2012; Prasad et al.,2012; Hameedet al.,2012) 、寄主植物( Dhawan et al.,2010; 黄芳等,2011; Guan et al.,2012; Muhammad et al.,2012;Kedar et al.,2012) 、饥饿( 郑婷等,2011) 等方面,关于光周期对棉花粉蚧生长发育和繁殖影响研究目前尚未见报道。作为影响昆虫生物学的重要因素,光周期可能对棉花粉蚧种群生存存在影响。生命表是研究昆虫种群动态的重要方法,也是研究害虫控制的重要方法之一( Harcourt,1969; 庞雄飞和梁广文,1995) 。应用生命表方法可研究阐明昆虫种群动态规律,预测种群消长趋势,系统评价防治措施效果,进而在此基础对害虫实施科学管理( 庞雄飞和梁广文,1995) 。本实验研究了光周期对棉花粉蚧实验种群生长发育和生命表参数的影响,可为深入了解该虫生物学和发生规律等提供参考信息,为该虫防治提供依据。
1、 材料与方法
1. 1 实验材料
供试虫源: 棉花粉蚧采于广州市珠江新城街道两旁扶桑 Hibiscus rosa sinensis,在室内以扶桑为寄主连续饲养、繁殖,取第 3 代备用。
供试植物: 将扶桑幼苗种植于华南农业大学昆虫学系温室,不施用任何化学药剂防治病虫,取新鲜、健康、干净的嫩叶若干备用。
人工气候箱: RXZ 型智能人工气候箱( 温度误差为 ±1℃,宁波江南仪器厂制造) 。试验设置 5 个光周期梯度: 8L∶ 16D,10L∶ 14D,12L∶ 12D,14L∶10D 和 16L∶ 8D,温度 27 ± 1℃ ,相对湿度为 70% ±5% ,光照强度 4 000 lx。
1. 2 试虫处理
在棉花粉蚧雌虫产若虫盛期,挑选刚产出的健康初孵若虫,用细毛笔小心将其接到盛有新鲜扶桑叶片的培养皿( Φ =9 cm,h =1. 2 cm) 中。每个光周期处理重复 3 次,每重复接虫 400 头。每天观察记录各处理棉花粉蚧蜕皮情况、各虫龄数量、羽化的雄虫数量、雌虫产仔数量及死亡虫数等,直至所有成虫产仔后生物学死亡为止。虽然棉花粉蚧雌成虫产仔后不再活动或取食,但是用细毛笔挑动虫体,仍然有轻微活动迹象,因为这样的雌成虫在生物学上已经失去存在价值,所以在本实验中视之为生物学死亡( Guan et al.,2012) 。
1. 3 生命表组建与分析
根据实验观察结果,参照 Birth( 1948) 及张孝羲和程遐年( 1997) 的方法组建棉花粉蚧繁殖特征生命表。该类生命表中各个参数具体含义如下:x: 棉花粉蚧的发育进程( d) ,以 1 d 为单位时间间隔; Lx: 逐日存活率,表示任何个体在 x 期间得以存活概率; Mx: 存活棉花粉蚧日平均产雌数,指在x 期间内平均每雌产雌数; 性比( ♀: ♂ ) 按照实际观察统计计算。根据棉花粉蚧不同光周期条件下繁殖特征生命表数据,应用 Harcourt ( 1969) 、赵志模和周新远( 1984) 、庞雄飞和梁广文( 1995) 等方法获得该虫实

1. 4 数据分析
对实验获得的棉花粉蚧各虫龄的虫数、发育历期、存活率、单雌产仔量等均采用 Duncan 氏新复极差法进行多重比较。由于 2 龄若虫末期棉花粉蚧雌雄分化,雌虫继续蜕皮进入 3 龄若虫期; 雄虫分泌丝状蜡质物包裹身体后进入预蛹期,计算时应将 3 龄若虫与 3 龄若虫和蛹区分开。
相关指标按以下含义和公式确定: 各虫龄虫数、雌雄成虫数是实验中实际观察到的虫体数量,1 龄若虫存活率 =2 龄若虫数量/1 龄若虫数量 ×100%;2 龄若虫存活率 = 3 龄若虫和雄虫蛹数量 /2 龄若虫数量 ×100%; 3 龄若虫存活率 = 雌成虫数量/3 龄若虫数量 ×100%; 3 龄若虫和蛹存活率 = ( 雌成虫 +雄成虫) /( 2 龄若虫数 ×2 龄若虫存活率) ×100%;单雌产仔量( 头) = 总产仔量/产仔雌虫数量。棉花粉蚧实验种群生命表中进入各发育历期虫数按照以下公式计算: 2 龄若虫数 = 初孵若虫数 ×1 龄若虫存活率; 3 龄若虫和蛹数量 =2 龄若虫数 ×2 龄若虫存活率; 成虫数量 =3 龄若虫和蛹数量 ×3 龄若虫和蛹存活率; 雌成虫数 = 成虫数量 × 雌成虫数/( 雌成虫数 + 雄成虫数) 。数据的分析与处理在 SAS9. 0 上进行,采用 Excel 软件进行绘图。
2、 结果与分析
2. 1 光周期对棉花粉蚧发育历期的影响
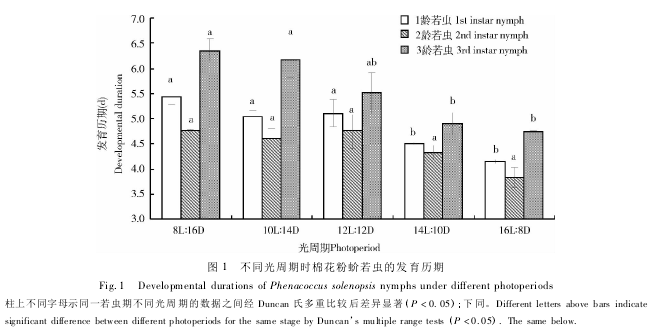
不同光周期处理后棉花粉蚧若虫的发育历期发生了变化,随着光照时间的延长,1 龄若虫、3 龄若虫和整个若虫期历期明显缩短( 图 1) 。14 和 16 h 光照时,1 龄若虫历期分别为 4. 51 和 4. 16 d,明显短于 12 h( 5. 11 d) 、10 h ( 5. 04 d) 和 8 h ( 5. 43 d) 光照时; 2 龄若虫各个光照长度下发育历期在 3. 83 ~4. 76 d,未发生明显变化( P = 0. 297) ; 3 龄若虫 16和 14 h 光照时历期为 4. 75 和 4. 90 d,显著短于 10h 光照时( 6. 17 d) 和 8 h 光照时( 6. 35 d) 。随着光照长度的增大,整个若虫发育历期逐渐缩短,8,10,12,14 和 16 h 时分别为 15. 54 ,15. 14,14. 55,14.36 和 12. 74 d。
2. 2 光周期对棉花粉蚧若虫存活的影响
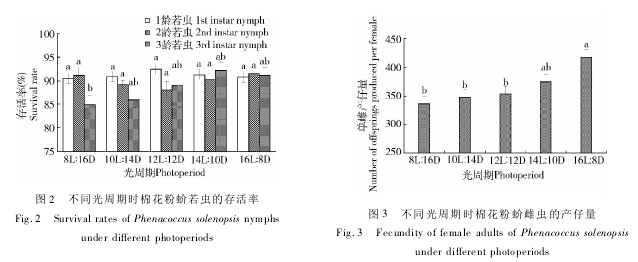
总体来讲,光周期对棉花粉蚧整个若虫期存活影响不明显( 图 2) 。8 ~ 16 h 光照时 1 龄存活率为90. 47% ~ 92. 36% ; 2 龄若虫为 87. 97% ~ 91. 49% ;短光照下3 龄若虫存活率出现下降现象,8 h 光照时若虫存活率为 84. 87%,明显低于 14 h 光照的92. 09% 。
2. 3 光周期对棉花粉蚧产仔能力的影响
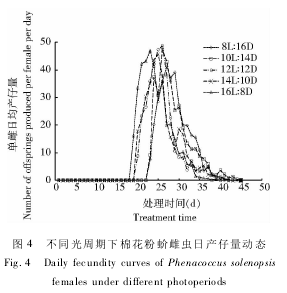
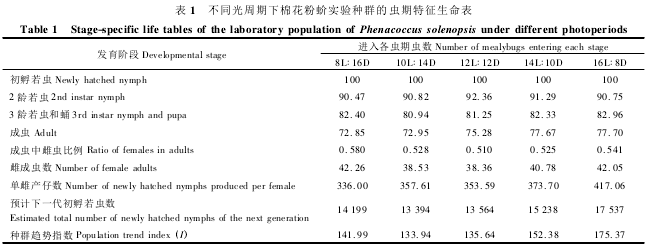
研究结果表明,长光照条件下棉花粉蚧产仔数量更大( 图 3) 。16 h 光照时单雌产仔数量最大,为417. 06 头; 12,10 和 8 h 光照时单雌产仔数量明显减少,分别为 353. 59,347. 61 和 336. 00 头; 14 h 光照时的产仔数量为 373. 70 头,介于 16 h 和 12 ~8 h两组之间。从雌成虫日产仔数量动态看,长光照处理较早进入产仔期,即产仔前期较短,如 16 h 光照处理产仔前期为18 d,14 和12 h 光照时产仔前期均为19 d,10 和8 h 光照处理均延长至22 d; 进入产仔期后一般 2 ~4 d 出现产仔盛期,此后持续一定时间后进入时间较长的盛末期( 图 4) 。
2. 4 光周期对棉花粉蚧种群繁殖的影响
2. 4. 1 棉花粉蚧实验种群繁殖特征生命表: 根据不同光周期下棉花粉蚧各虫期存活率和成虫繁殖力观察结果,组建了该虫实验种群的虫期特征生命表( 表 1) 。表 1 中的起始初孵若虫数 100 为假定数值,实际起始虫数超过 1 200 头,各发育阶段的存活数为各发育阶段虫数占原始初孵若虫数的百分比乘 100。与短光照相比,长光照条件的种群趋势指数明显较大,光照 16 h 时为 175. 37; 14 h光照时较大,为152. 38; 8,12和10h光照时分别降低至 141. 99,135. 64 和 133. 94。本研究中无论哪个光周期处理,棉花粉蚧下一代的数量均将显著增大到当代的 100 余倍,这表明该虫种群增殖潜能巨大,具备迅速暴发成灾的生物学特性,而长光照更有利这种害虫种群的增长。
2. 4. 2 棉花粉蚧实验种群参数: 组建了不同光周期下棉花粉蚧繁殖特征生命表( 表 2) ,并根据时间存活率及成虫繁殖力分析获得了该虫实验种群生命表参数( 表 3) 。由该表可知,随着光照时间加长,棉花粉蚧种群的净增值率、内禀增长率、周限增长率这 3 个参数均表现出逐渐增大的趋势。
8 ,10 ,12 ,14 和 16 h 光照时净增值率由 88. 57和 86. 85 逐渐增大至 100. 77,124. 16 和 126. 86;内禀增长率由 0. 1616 增大至 0. 2066; 周限增长率由 1. 1754 增大至 1. 2294。与以上 3 个参数相比,种群倍增时间则表现为相反的趋势,即随着光照时加长而逐渐缩短,8 和 16 h 光照时由 4. 29 d 降低至3. 36 d。
4、 讨论
光周期对昆虫的生长发育与繁殖有着重要作用( Roditakis et al.,2001; Keena et al.,2012; Kollberget al.,2013; Zerbino et al.,2013) 。许多昆虫对光敏感的时期主要集中在若虫/幼虫期。Dole?al 等( 2007 ) 报道在短光照处理下,马铃薯甲虫Leptinotarsa decemlineata ( Say) 幼虫发育速率加快,食物消耗减少,但成虫不产卵。赵洪霞等( 2011) 发现在较长光照和较短光照条件下,绿盲蝽 Lyguslucorum Meyer-Diir 若虫的发育速率加快,雌成虫产卵期也缩短,产卵量减少。本研究发现光周期对棉花粉蚧的发育历期和生殖的影响是显著的。在 5 个光周期条件下较短光照( 8 h) 处理棉花粉蚧若虫发育历期最长,发育速率最慢,且雌成虫产仔前期较长,产仔量最小; 较长光照( 16 h) 处理若虫发育历期最短,发育速率最快,且雌成虫产仔前期较短产仔量最大。这说明长光照是有利于棉花粉蚧发育和生殖。根据中国棉花粉蚧的潜在适生区域预测的结果( 马骏等,2011) 和光周期资料,给出该虫适生区域的光周期变化范围约为 8L∶ 16D ~ 16L∶ 8D。因此,在研究棉花粉蚧种群暴发规律时除了包括温度、湿度、降水、寄主、天敌等因素外,可能还需要考虑光照时间这个因素,这样可能更全面和准确一些。
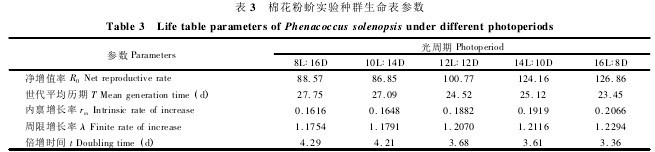
生命表方法广泛用于研究、分析各种因子对昆虫种群数量变动的影响。一份完整的生命表可以直观地反映种群数量动态的某些特征( 例如发育历期、存活率、产雌率等) 。对生命表信息进行分析,寻求种群数量变动的内在规律,研究害虫发生规律,并作出预测,同时找出昆虫生活史中最薄弱的时期,以便制定适合的防治措施。本研究组建了棉花粉蚧实验种群虫期特征生命表、繁殖力特征生命表,并给出了实验种群生命表参数。种群的内禀增长率( rm) 是反映种群特性的一个重要指标,它综合了昆虫的存活率、发育速率、产卵速率以及年龄组配、产卵量等因素,因此能更精确地表述种群的变化情况( 戈峰,2008) 。在光周期为8L∶16D 时棉花粉蚧需要 27.75 d 完成一个世代,而在没有其他外界因子影响下一个世代后数量将增长到141.99 倍; 16L∶ 8D 时该虫 23. 45 d 完成一个世代后数量将增长至175.37 倍。排除其他因子的作用,该种群趋势指数很大,表明该虫种群具备迅速增长的潜能。综上所述,本研究结果表明光周期对棉花粉蚧的生长、繁殖和种群增长影响显著。





