
烟粉虱 Bemisia tabaci 是一种世界性入侵害虫,不但可以直接为害多种蔬菜等作物,并且能够通过取食传播番茄黄化曲叶病毒 ( Tomato yellow leafcurl virus,TYLCV) 等病原微生物 ( Oliveira et al. ,2001) ,造成更大的经济损失。烟粉虱上世纪 90年代在我国开始暴发,在此后的 20 年间造成了极为严重的危害。尽管通过使用化学杀虫剂可以暂时减少烟粉虱的虫口密度,但是同时加速了烟粉虱在种下生物型的分化,以及高抗药性的形成( Prabhaker et al. ,1985) .自21 世纪初开始,人们尝试应用各种天敌昆虫对烟粉虱进行生物防治。
其中寄生性天敌中恩蚜小蜂属的种类最多,如丽蚜小蜂 Encarsia formosa ( Gahan) ( Hymenoptera:
Aphelinidae) 已成功应用于温室蔬菜上烟粉虱的生物防治 ( Hoddle et al. ,1998) .此外,浅黄恩蚜小蜂Encarsia sophia ( Girault Dodd) 对烟粉虱的防治也是一种有很好的应用前景的寄生蜂 ( 李元喜等,2008) .
大量研究表明,单纯利用某种寄生蜂建立生物防治种群,其防治效率会随着世代数增加而线性降低 ( Barbosa,1998) .而将不同天敌昆虫混合释放,不仅可以间接增加群落的稳定性,而且在提升相同食物链内生物多样性的同时,减少天敌种群的自然衰退 ( Heimpel & Mills,2008) .Malo等 ( 2012) 进行了大量同资源种团 ( Intra - guildagents) 的研究,但是关于寄生蜂之间以寄主为媒介的竞争关系研究及其对烟粉虱防控效果的评价却鲜见报道。
本文选择蚜小蜂科恩蚜小蜂属的专性寄生蜂浅黄恩蚜小蜂作为研究对象,在室内环境下对其单独和其近似生态位的丽蚜小蜂混合释放时与烟粉虱寄主密度的伴随关系进行研究,并通过拟合Holling 功能反应方程,评价浅黄恩蚜小蜂在不同温度下单独及混合应用的寄生潜力。
1 材料与方法
1. 1 供试虫源
本实验中所用浅黄恩蚜小蜂、丽蚜小蜂均为北京市农林科学院植物保护环境保护研究所室内定殖种群。两种寄生蜂分别隔离饲养于不同的养虫笼 ( 铝合金 +80 目纱网制,45. 0 cm × 50. 0 cm× 50. 0 cm) 内,控制虫口密度为 50 - 70 头 / 笼。
寄生蜂定殖及试验所用寄主 B 型烟粉虱采集自北京市昌平区有机蔬菜温室内,室内以番茄 ( 品种:一品红) 作为寄主植物扩繁。在寄生蜂定殖过程中,每 12 h 向养虫笼内补充带有足量烟粉虱的番茄叶片。实验种群扩繁环境条件由室内温湿度控制系统调控,各参数设置为: 温度 25℃,湿度65% ,光照周期 = 16 L∶8 D,光照强度 = 800 lx.
1. 2 不同温度下浅黄恩蚜小蜂对不同密度烟粉虱三龄若虫的寄生
取带有烟粉虱 3 龄若虫的番茄叶片,分别设密度为 2,4,6,8,10,12,14,16 头处理,将多余的烟粉虱用软尖笔剔除。分别将各处理带有烟粉虱若虫的番茄叶片和一头已交配的浅黄恩蚜小蜂雌成虫同时放于培养皿 ( D =9. 0 cm) 中,任其自由寄生 24 h.后将浅黄恩蚜小蜂吸除,利用分子生物学快速检测方法 ( Zhang et al. ,2013) 对不同密度下烟粉虱的寄生量进行检查,并分析统计。
本试验在光照培养箱 ( Sanyo - MH351) 内完成,温度处理分别设置为 15,20,25,30,35℃,其余各项环境参数设置同 1. 1.各试验处理重复 10 次。
1. 3 不同温度下浅黄恩蚜小蜂与丽蚜小蜂混合对不同密度烟粉虱三龄若虫的寄生量
按照上述步骤准备带有不同密度 ( 4,8,12,16,20,24,28,32 头) 烟粉虱的番茄叶片,与一头新交配的浅黄恩蚜小蜂雌成虫以及一头丽蚜小蜂雌成虫同时放置于同样规格的培养皿中。其余试验方法与环境参数同 1. 2.
1. 4 功能反应方程及搜寻效应系数拟合
依据前述试验结果,分别获得浅黄恩蚜小蜂独立以及与丽蚜小蜂混合存在时,在不同温度条件下对不同密度烟粉虱三龄若虫的寄生量,以Holling Ⅱ型功能反应模型为对象进行拟合:【1】
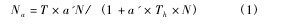
式中,Th为处置时间; T 为试验完成的总时间 ( 本试验中为 1 d) ; a‘为捕食者对猎物的瞬间攻击率; N 为猎物密度,Na为被寄生的猎物数量。
并通过上式所得参数计算出理论最大寄生量Na max = 1 / Th.由拟合 Holling Ⅱ型功能反应方程所得各参数按照下式计算不同处理下浅黄恩蚜小蜂对烟粉虱三龄若虫的搜寻效应 S:【2】
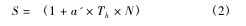
式中各参数意义同式 ( 1) 所标注内容。
2 结果与分析
2. 1 不同温度条件下浅黄恩蚜小蜂对烟粉虱 3 龄若虫的寄生功能反应
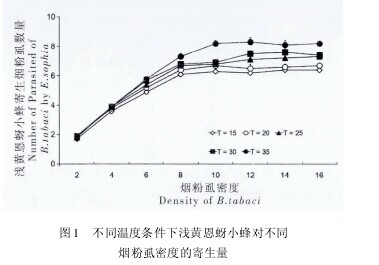
如图 1 所示,已交配浅黄恩蚜小蜂在本试验所设置的各个温度条件下,其寄生量均受寄主烟粉虱密度的显着影响 ( F15= 21. 371,P15< 0. 01;F20= 27. 448,P20< 0. 01; F25= 19. 638,P25< 0. 01;F30= 26. 107,P30< 0. 01; F35= 22. 535,P35< 0. 01;所有 d. f. = 7,72) .在同一寄主密度下,浅黄恩蚜小蜂对烟粉虱的寄生量随着温度的上升而增加。
按照式 1 拟合 Holling Ⅱ型功能反应方程,结果显示,本试验中各温度下浅黄恩蚜小蜂对烟粉虱的寄生均表现为寄主密度制约型,与 Holling Ⅱ型功能反应方程高度拟合 ( R >0. 9) .经 X2检验,表1中不同温度下浅黄恩蚜小蜂的实际寄生量与各密度下按照所拟合方程计算的理论值差异均不显着( P >0. 05) .表 1 说明,浅黄恩蚜小蜂在寄生烟粉虱时的处置时间,会随着环境温度的上升而减少;对烟粉虱的瞬间攻击率会随着温度上升而增加。【表1】
当寄主密度趋于无穷时,参数 1/Th可以反应出寄生物的理论最大寄生量。在本研究中,已交配的浅黄恩蚜小蜂对烟粉虱三龄若虫的最大理论寄生量也同样随着环境温度的上升而增加。【图1】
利用表 1 中所得功能反应模型参数 a’和 Th,按照式 2 拟合得到了浅黄恩蚜小蜂在不同温度下对烟粉虱三龄若虫的搜寻效应系数 S ( 表 2) .在相同温度条件下,浅黄恩蚜小蜂对于寄主烟粉虱的搜索效应均随着寄主密度的上升而增加。而在任意寄主密度下,浅黄恩蚜小蜂对于烟粉虱三龄若虫的搜索效率均随着环境温度的增加而上升。【表2】
2. 2 不同温度条件下浅黄恩蚜小蜂与丽蚜小蜂混合处理对烟粉虱 3 龄若虫的寄生功能反应
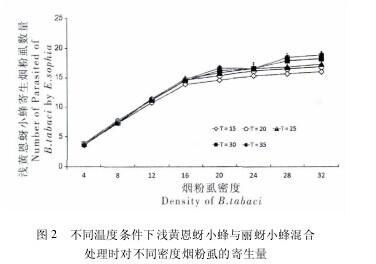
在与丽蚜小蜂同时存在的情况下,已交配的浅黄恩蚜小蜂在各温度下对烟粉虱的寄生率同样随寄主密 度 的 上 升 而 显 着 增 加 ( 图 2. F15=39. 743,P15< 0. 01; F20= 42. 556,P20< 0. 01; F25= 41. 534,P25< 0. 01; F30= 39. 852,P30< 0. 01;F35= 29. 844,P35< 0. 01; 所有 d. f. = 7,72) .在同一寄主密度下,浅黄恩蚜小蜂对烟粉虱的寄生量也会随着温度的上升而增加。混合种群内浅黄恩蚜小蜂对烟粉虱密度的寄生功能反应拟合结果如表 3 所示。当两种寄生蜂混合时,各温度下浅黄恩蚜小蜂对烟粉虱的寄生同样表现为寄主密度制约型,与 Holling Ⅱ型功能反应方程高度拟合( R >0. 9) .经 X2检验,表 3 中不同温度下浅黄恩蚜小蜂的实际寄生量与各密度下按照所拟合方程计算的理论值差异均不显着 ( P > 0. 05) .与浅黄恩蚜小蜂单独处理情况相同,浅黄恩蚜小蜂在寄生烟粉虱时的处置时间,也随着环境温度的上升而减少。与此同时,其对烟粉虱的瞬间攻击率会随着温度上升而增加。浅黄恩蚜小蜂对烟粉虱三龄若虫的最大理论寄生量也同样随着环境温度的上升而增加。【图2】
按照表 3 中所示各项功能反应参数计算出不同温度、不同寄主密度下,浅黄恩蚜小蜂在与丽蚜小蜂同时存在时的搜索效应系数 ( 表 4) .与浅黄恩蚜小蜂单独寄生时结果相似,各温度下,混合寄生蜂中下浅黄恩蚜小蜂的搜索效应随寄主密度的增加而上升,同一寄主密度下,其寄生效率同样随温度的上升而增加。【表3.表4】
3 结论与讨论
通过拟合功能反应模型,比较功能反应参数可以直观的了解高营养级的天敌昆虫对靶标害虫的控制潜能。其中,瞬间攻击率 a‘以及单位靶标处置时间 Th可以清晰的反应出天敌昆虫在整个控害过程中的行为策略及控害效率 ( Holling,1966) .在本文中,浅黄恩蚜小蜂单独寄生烟粉虱和与丽蚜小蜂同时寄生烟粉虱两个处理,其寄生量均为密度制约型,即寄生量随寄主密度的上升而上升,符合 Holling Ⅱ型功能反应模型。
在本试验中,浅黄恩蚜小蜂的寄生能力受环境温度变化影响显着。其寄生效率随温度的上升而增加。在高温下其瞬间攻击率最高,且寄生处置时间最短,而此时理论最大寄生量最高。大量类似研究表明,寄生蜂的寄生效能随温度变化而波动显着。施祖华和刘树生 ( 1999) 研究发现菜蛾绒 茧 蜂 Cotesia plutellae 在 高 温 下 对 小 菜 蛾Plutella xylostella 的寄生效率要显着高于低温环境,其产卵寄生的雌蜂比例随温度的升高而增加。同样,当斜纹夜蛾侧沟茧蜂 Microplitis sp. 寄生斜纹夜蛾 Spodoptera litura 幼虫时,其瞬间攻击率随温度上升而增加,而寄主的处置时间相应减少 ( 蒋杰贤等,2002) .这些研究表明,温度是制约寄生蜂寄生行为的重要因子。较高的瞬间攻击率及较短的寄生处置时间说明寄生蜂的寄生行为随温度上升而显着增加。施祖华等 ( 1999) 认为寄生蜂寄生时,保持静止状态的比例会显着影响功能反应参数的变化。在本试验中,浅黄恩蚜小蜂在低温状态下有很大的比例是处于静止状态的,这样的时间区段不能作为搜索和处置时间来归类,因此这对于温度和寄生蜂寄生效率关系的影响在后续研究中同样值得考虑。此外,寄生蜂在高温下的优越表现与烟粉虱所处的温室生态系统中高温环境相符合,因此也从一定程度上表现出其在温室生态系统中防控烟粉虱的潜能。
一直以来,人们尝试通过混合释放天敌昆虫来增强生物防治功效 ( Letourneau et al. ,2009) .但是室内研究表明不同天敌昆虫之间由于存在生态重叠以及生态位竞争,共同释放时所产生的增效作用十分有限。Hagler 等 ( 2004) 发现多种捕食性天敌混合释放时,天敌之间会自动调节,继而分食不同发育阶段的烟粉虱,从而达到一种相对平衡的状态。Collier & Hunter ( 2001) 研究发现浅黄恩蚜小蜂与另一种蚜小蜂 Eretmocerus eremicus混合释放时,两者不但相互影响对方种群的交配繁殖效率,并且还进一步影响各自对温室粉虱Trialeurodes vaporariorum 的寄生效率以及取食情况。
而在本研究中当浅黄恩蚜小蜂与丽蚜小蜂混合释放时,浅黄恩蚜小蜂的寄生效率并没有降低,其对寄主的瞬间攻击率较单独存在时呈现上升的趋势,且其对单头寄主的处置时间明显减少。可见与相近生态位寄生蜂混合释放后,浅黄恩蚜小蜂对于烟粉虱的潜在防控能力并没有得到抑制,相反其为了减少竞争者的影响,主动提高了对寄主的寄生能力。而这一变化,会直接提高其对烟粉虱的防治效果。在本试验中,我们仅仅在室内考虑了浅黄恩蚜小蜂与丽蚜小蜂等量组合的效果。
而在田间实际应用中,不同的配比组合是否会进一步增强浅黄恩蚜小蜂的生物防治功效,还需要进一步深入研究。





