摘 要
利用体外标记法对江苏南通、辛庄、谢桥、大义和白茆克氏原螯虾交配情况进行观察,共记录到 116 对交配虾。其中,异地交配的克氏原螯虾为 93 对,占总数的 80.17%;同地交配的为 23 对,占总数的 19.83%。不同地区的克氏原螯虾在选择交配对象时没有表现出对某个地区的虾特别喜好。克氏原螯虾更趋于与自身体重和体长相近的虾进行交配,体重差值在 5g 以下与体长差值在 0-1cm 间的交配对数分别占总数的 56.90%和 86.21%。结果表明:克氏原螯虾在性选择时存在一定机制指导其避免近亲交配,选择异地交配。在个体大小的选择时趋向于选择和自身相当的虾。
关键词:克氏原螯虾 性选择 多样性
The selective mates analysis of the crayfish Procambarus clarkii
Abstract
Using in vitro labeling to observe the crayfish Procambarus clarkii mating situation ofNantong,Jiangsu,Xinzhuang,Xieqiao,Dayi and Baimao,which were recorded 116 pairs of matingcrayfishes.Among them,there were 93 pairs of remote mating crayfishes,accounting for80.17%;the same place for 23 pairs of mating,accounting for 19.83%.Different regions ofcrayfish in the choice of a mate did not demonstrate pecial preferences to a region of the crayfish.The crayfish Procambarus clarkii tends to be more similar to their own body weight and bodylength of crayfish to mate,the difference between body weight and body length in the following5g difference between the 0-1cm mating on the total number of 56.90%,and 86.21%.The resultsshowed that:there is a certain mechanism of choice in the crayfish Procambarus clarkii to guideits avoid inbreeding,but distant objects affinity selection. And the size of the individual to makechoices and choose their own considerable crayfishes.
Key words: Procambarus clarkii; sexual selection; diversity
目 录
1.引言
1.1 克氏原螯虾简介
1.2 克氏原螯虾的繁殖生物学
1.2.1 虾的怀卵量
1.2.2 繁殖时间
1.2.3 亲虾雌雄鉴别
1.2.4 雌雄性比
1.2.5 卵巢发育
1.2.6 精巢发育
1.2.7 交配行为
1.3 选择性交配研究进展
1.3.1 与个体大小有关的性选择行为
1.3.2 与“婚礼食物”有关的性选择行为
1.3.3 精子竞争
1.3.4 与鸣声有关的性选择行为
1.3.5 影响雌、雄选择的因素
2.实验材料与方法
2.1 实验材料
2.2 实验方法
2.2.1 试验虾的处理及观察
2.2.2 养殖条件控制
2.2.3 实验数据处理
3.实验结果
3.1 克氏原螯虾异地交配与同地交配观察
3.2 不同地区克氏原螯虾地域选择性交配观察
3.4 克原螯虾交配对体长大小的选择性
3.4 克氏原螯虾交配对体重大小的选择
3.5 克氏原螯虾雌、雄选择性
4.讨论
4.1 克氏原螯虾对交配对象地域的选择
4.2 克氏原螯虾对交配对象体长大小的选择
4.3 克氏原螯虾对交配对象体重大小的选择
5.参考文献
6.致谢
1.引言
1.1 克氏原螯虾简介
克氏原螯虾(Procambarus clarkii)俗称“小龙虾”,隶属于节肢动物门(Arthropoda),甲壳纲(Crustacea),十足目(Orgerdecapoda),爬行亚目(Reptantia),螯虾科(Cambaridae),原螯虾属(Procambarus)。原产于北美洲,自日本引入我国后,种群发展迅速,并且在很多水域已形成优势群体,成为重要的水产资源[1]。克氏原螯虾是我国的外来物种,80 多年来已遍布全国,在分布广泛的地区其种群遗传多样性仍处于较高水平,只是在少数群体较小的地区遗传多样性有所降低,不过仍处于中等水平[2-3]。克氏原螯虾在发展中一直都能保持较高的遗传多样性,研究其保持多样性的机制对克氏原螯虾的种质保护具有重要意义。
性选择是指雌雄个体间根据第二性征进行选择性交配的一种婚配现象,广泛存在于自然界[4]。雌雄形体外貌特征差异、体色和形体大小等都对性选择的方向产生影响。目前,陆生生物性选择研究较多,水生生物的相关研究主要集中在鱼类[5-6],海胆[7]、牡蛎[8]、蟹类和福寿螺[9-10]也有少量报道,但克氏原螯虾的研究还未有。本文对克氏原螯虾的性选择进行了初步研究,以期为克氏原螯虾遗传多样性研究提供一定的科学依据。
1.2 克氏原螯虾的繁殖生物学
1.2.1 虾的怀卵量
王顺昌(2003)报道,克氏原螯虾的怀卵量一般在100粒~700粒,平均为300粒。王福刚等(1995)报道,不同年龄的红螯螯虾怀卵量也不一样,初次产卵且体长小于10cm的,怀卵量约为200粒~400粒,体长超过10cm的,怀卵量约为600粒;同一体长超过10cm的亲虾,第二次怀卵量可达到1000粒。青虾的相对怀卵量在600-700粒/g之间(潘华等,2002)。
壳氏原螯虾在性成熟后每次繁殖前雌虾都会蜕一次皮(唐鑫生,2001),罗氏沼虾、青虾也有这种现象(崔淑贞等,1995;潘华等,2002),也称为生殖蜕皮。
1.2.2 繁殖时间
红螯螯虾的产卵高蜂在5月~7月(Hsin-Shemg et al.,1995)。但由于地区不同,产卵时间可能也有不同,在浙江北部,该虾的产卵高峰期为4月~6月 (顾志敏等,2003)。在厦门、山东其繁殖期为5月~10月份,盛期出现在6月~8月份(谢开恩等,1995;初世伟等,2003)。
1.2.3 亲虾雌雄鉴别
王顺量(2003)报道,雄虾腹部第一游泳肢特化为交和刺,而雌虾第一游泳肢特化为纳精囊,唐鑫生(2001)报道,同龄亲虾个体,雌虾比雄虾大;体长相近的亲虾,雄性螯肢比雌性的粗大,且雄性螯肢的腕节和掌节上的棘突长而明显。短螯螯虾的雌雄特征和克氏原螯虾的相近,位于雄虾第五对步足基部,具有一对球形的、细长而带硬棘的棒状生殖突起,交配时精荚从这里排出。雌虾在第三对步足基部有一对生殖孔,成熟卵从这里排出(赵云龙等,2000)。
1.2.4 雌雄性比
Carmona 等(2004)报道,研究虾的性别搭配比例有助于虾的成功繁殖,但目前这种比例还不太清楚。不同的试验者得出的性别比是不一样的。对于克氏原螯虾,郑楚龙(1995)则认为雄雌比例应为 1:4~6。日本对虾雌雄比例为 1:1。青虾的雌雄比在繁殖初期为1.107:1,在产卵盛期为 1.439:1(潘华等,2002)。Assaf 等(2000)报道一种红螯虾繁殖时雌雄比例达到 5:1。Christopher 等(1999)研究表明不同地区的一种淡水龙虾的两个亚种的雌雄搭配比例是不同的,但这种不同搭配的机制还有待进一步研究。
1.2.5 卵巢发育
卵巢发育的好坏与虾的质量、个体大小、生活环境、培育条件、年龄有着密切关系。
王顺昌(2003)根据克氏原螯虾的性腺发育时卵巢的颜色的变化来判断性腺成熟程度,可把卵巢发育分为苍白、黄色、橙色、棕色(茶色)和深棕色(豆沙色)等阶段,颜色越深性腺发育就越好;Oscar 等(1995)对克氏原螯虾的卵巢发育的分期和王顺昌的差不多,其中苍白色是未成熟幼虾的性腺,细小,需数月方可达到成熟;橙色是基本成熟的卵巢,交配后需 3 个月左右可以排卵;茶色和棕黑色是成熟的卵巢。朱崇梅(2000)认为克氏原螫虾分期成熟,属多次分批产卵型;王克行(1996)也认为克氏原螯虾有两个产卵高峰期,分别在春季的 4 月~5 月和秋季的 9 月~10 月。Claudia 等(2004)报道,在 16℃,21℃,26℃三种温度当中,随温度的升高卵巢发育越好,且前种温度与后两种温度的发育存在显着差异,但后两种温度之间不存在。关于克氏原螫虾的性腺发育机制,Rachakonda 等(1996,1997)研究表明,蛋氨酸—脑啡肽和纳洛酮都能对克氏原螯虾体内的胸神经中枢作用,能调节释放 GSH,促进卵巢的发育。Antonio R(2000) 摘除眼柄的方法可以很好的促进其卵巢的发育。Rachakonda&Sarojim 等(1995)研究萘与克氏原螯虾卵巢发育之间的关系时发现,用 lOg/m3 萘浸泡了 15d 的雌虾卵巢中的卵黄前期和卵黄期存在退化,但对于浸泡的未成熟的雌虾再返回清水中时,其卵巢发育会继续。
1.2.6 精巢发育
关于十足的甲壳动物精巢发育方面尤其是精子发生方面的研究,国内外已经,有很多报道。如堵南山等(1988)、王义磊等(1996,1997,1998)、秋高蜂等(1997),王风玉等(1998)、杨万喜等(1998,1999)、Jeapersen(1979。1983)、Pochon,J(1983)、Nicotra,a.等(1986,1985)。而关于虾类精巢的组织学分期,洪水根等(1998,1999)、罗宇良等(1999)都有没统一的划分标建,罗宇良等将红螯螯虾的精巢发育分为:未发育期、发育期、成熟期和休整期;而日本学者 T.Hamano(1990)则把口虾咕的精巢分为 5 个期,他将该虾的成熟期分为成熟前期和成熟期。而且这些分期都是在借鉴别人研究基础上结合作者的研究对象的特点再进行划分的。
1.2.7 交配行为
邱高峰等(1995)研究表明,克氏原螯虾的交配行为过程可分为交配前期和交配期。
在交配前期,雄虾表现出特有的交配前期行为;交配期包括钳夹、翻转、横跨、交尾和分离五个阶段。雌虾被雄虾钳住后表现顺从行为;而在对照组中的雌虾被同性钳住质却不表现顺从行为,雄虾之间的接触导致进攻行为。交配时雄虾横跨的步足均为右第 5 步足;雌虾在交配前匆需生殖蜕皮,交配的成败与雄虾接近雌虾的行为方氏有关,且螯足较大的雄性个体交配成功率明显较高。红螯螯虾的交配行为和克氏原螯虾的相似;但是,红螯螯虾交配条件是必须是雌虾刚蜕皮的 24h 内交配(赵云龙等,2000)。虾交配持续的时间为 5min~6min(唐鑫生,2001),邱高峰等(1995)通过观察发现该虾交配持续的时间为几小时,甚至十几时。
1.3 选择性交配研究进展
目前尚无克氏原螯虾交配选择性的研究,主要借鉴国内外对昆虫、蟹类、鸟类和福寿螺等的研究。
1.3.1 与个体大小有关的性选择行为
在雄性的相互竞争中,个体大的力量更加大,因此交配的成功率就比较高。Silja etal(1998)对叶甲(galerucella nymphaeae)雌性的交配选择做了研究。结果表明,当雌性叶甲与体型大小不同的 2 只雄性同时相处的时候,则雌性更喜欢与前胸背板较宽的雄性进行交配。这就表明这种叶甲雄性与雄性之间在交配竞争方面,个体大小是最主要的因素。
与个体大小相关的交配行为中,往往呈现出一种现象,即个体大的雌性与个体小的雄性进行交配,而个体小的雌性则与个体大的雄性进行交配。个体大的雄性往往更加灵活。因此,在寻找配偶以及寻找交配场所方面占有优势,交配成功率相对比较高。RobertB(2002)对一种沫蝉(Cercopis sanguinolenta)进行性选择实验。结果表明,雌性优先与个体较大,灵活性比较强的雄性个体交配。
1.3.2 与“婚礼食物”有关的性选择行为
精子保护假说认为婚礼食物作为雄性取悦雌性的“彩礼”,雄性送给雌性“礼物”以获得受精权。在雌性取食精包的时候,精子趁机进入雌虫体内,卵子排出时完成受精。
王孟卿等(2005)及长有德等(2002)认为雄性提供“彩礼”的能力通常与个体大小呈正相关,个体较大的雄性往往会在雄雄竞争获胜或受到雌虫的“青睐”。
1.3.3 精子竞争
许多昆虫有多次交配行为,交配后不同雄虫个体的精子被较长时间地贮存在雌虫的受精囊内。这样,雌虫卵子受精时,就会发生不同雄虫的精子竞争(sperm competition)。
精子竞争也是雌虫一种主要而有效的性选择方氏,也是雄性竞争的最后形氏[11]。可分为:
(1)最先雄虫精子优先(first male sperm precedence)(2)最后雄虫精子优先(last malesperm precedence);(3)无先雄虫精子优先(no male sperm precedence) 。大多数的昆虫表现为最后雄虫精子优先,Gwynne[12]统计了 37 种昆虫精子的竞争,发现72.9%的种类为最后雄虫精子优先,18.9%的种类为最先雄虫精子优先,8.2%的种类无雄虫精子优先。
1.3.4 与鸣声有关的性选择行为
昆虫的声通讯行为主要是通过鸣声来实现的,鸣声在种内个体间的召唤、求偶、攻击和报警等方面起着非常重要的作用。因此,昆虫通过鸣声来选择配偶也是比较常见的对一些能够发声昆虫的性选择行为进行了研究。结果表明,这些昆虫的雌性会优先与鸣声能量高的雄性进行交配[13]。目前有人提出一种假设即“求偶鸣曲信息传递”,该假设认为:求偶鸣曲作为吸引异性的交配信号,可表达和传递身体质量优劣等相关信息,异性通过鸣曲判断对方优劣,最后决策接受还是拒绝与之交配[15]。
1.3.5 影响雌、雄选择的因素
分为两种:雌选雄、雄选雌。两种选择在特定情况下有可能转变。性选择作用的强度和方向依赖于两性生殖投资的差异和获得配偶的难易程度。当两性的投资的差异大体相等时,如在单配性鸟类中雄鸟和雌鸟共同喂养雏鸟,性选择作用的强度就比较弱。两性的生殖投资差异很大时,通常会发生激烈的配偶竞争,即投资少的一性(通常是雄性)为争夺投资多的一性而展开竞争(尚玉昌,1998)。种群密度影响可获得食物在整个种群中的分布,从而间接影响雄性提供“彩礼”的能力。摩门螽斯(Anabrus simplex)在低密度下表现正常的角色,而在高密度下表现性角色逆转现象。表现为雌性聚集在一起,雄性在其中选择配偶(Gwynne,1984)。
2.实验材料与方法
2.1 实验材料
实验用克氏原螯虾分别取样于江苏南通,辛庄,谢桥,大义,白茆。实验虾在实验室内按不同地区雌雄分开暂养于玻璃缸中,放入假草作为隐蔽物,以小鱼为饵料,暂养期间每2天换一次水,暂养缸充氧且控制温度,暂养一周以上,使其适应实验环境,以便取样。
2.2 实验方法
2.2.1 试验虾的处理及观察
随机选取体色好,健康活泼,肢体健全的成体虾(每个地方雌雄各 10 只)用不同颜色的油彩笔在每只虾的头胸甲上做好标记(表 1),然后放入同一个玻璃缸,设置三个平行。每天从早上 7:00 到晚上 23:00 每隔 2-3 小时观察交配情况,发现有抱对交配的虾立即捞出,放入单独的桶中,记录交配虾头胸甲上油彩笔的颜色(地区),分别称重,测量体长。5 天加一次虾,让缸中始终保持 100 只虾,观察时间一个月。另随机选取体色好,健康活泼,肢体健全的成体虾(雌雄各 3 只)用油彩笔在胸甲上作好雌雄记号并放入同一个水盆中,水盆中间用挡板隔开,仅在中间开一小孔,供单只虾通过,一边放置雄虾,另一边放置雌虾,设置 3 个平行。每天从早上 7:00 到晚上 23:00 每隔 3-4 小时观察并记录盆内虾分部情况,观察时间一个月。
2.2.2 养殖条件控制
实验过程中以小鱼为饵料,每天晚上投喂一次;实验缸及盆中放入两个充氧头使溶氧达到 5mg/L,并用控温仪控制水温为 27oC;每 2 天换水一次,换水时用虹吸管将缸底及盆底的残饵和粪便吸尽,再加入经曝气的新水。
2.2.3 实验数据处理
实验数据记录在 EXCEL 表上,采用 Origin 软件分析检验实验所得数据,P>0.05 表示差异不显着,P<0.05 表示差异显着,P<0.01 表示差异高度显着。
3.实验结果
3.1 克氏原螯虾异地交配与同地交配观察
实验期间共观察到了 116 对虾进行交配,在交配的 116 对中,异地交配的为 93 对,占总数的 80.17%(P<0.01);同地交配的为 23 对,占总数的 19.83%(图 1)。实验结果表明:克氏原螯虾在交配时有一定的地域选择性,大多数克氏原螯虾趋向选择异地虾交配。
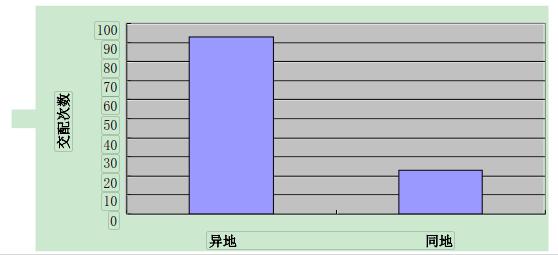
图 1 异地交配与同地交配次数
3.2 不同地区克氏原螯虾地域选择性交配观察
实验中与辛庄虾交配的有 29 只,辛庄虾交配的选择没有特别喜爱的地区,但大多是与异地交配,和同一地区的交配较少(图 2-1)。与谢桥虾交配的有 44 只,大义的比例最大(37%),其次是白茆和南通,均为 20%(图 2-2)。与南通虾交配的有 23 只,其中辛庄和谢桥占绝大部分比例(35%和 39%)(图 2-3)。与大义虾交配的有 66 只,大义的虾趋向于与白茆虾进行交配(41%),与谢桥虾次之(24%)(图 2-4)。与白茆虾交配的有 46 只,白茆虾交配时倾向于选择大义的虾(58%)(图 2-5)。而用 origin 软件进行方差分析表明,在 0.05 水平上,各地区虾对不同地域虾的选择并无显着差异,即虾的地域选择是随机的。
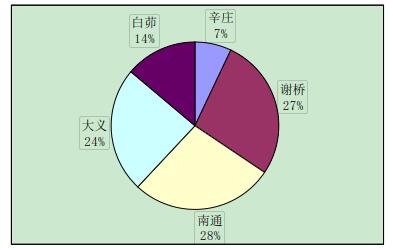
图 2-1 辛庄克氏原螯虾对不同地区克氏原螯虾的交配选择性
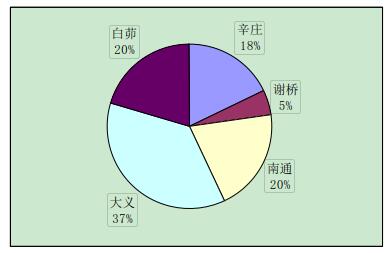
图 2-2 谢桥克氏原螯虾对不同地区克氏原螯虾的交配选择性
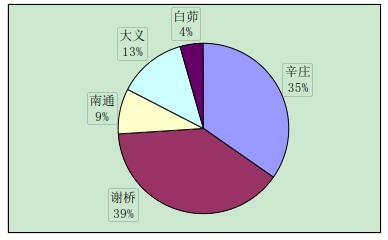
图 2-3 南通克氏原螯虾对不同地区克氏原螯虾的交配选择性
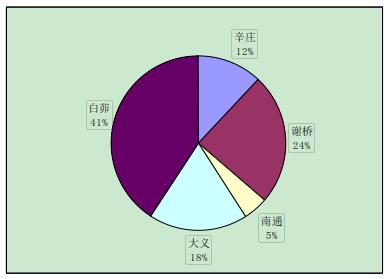
图 2-4 大义克氏原螯虾对不同地区克氏原螯虾的交配选择性
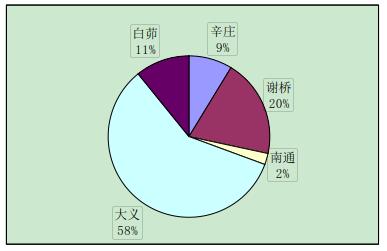
图 2-5 白茆克氏原螯虾对不同地区克氏原螯虾的交配选择性
3.3 克原螯虾交配对体长大小的选择性
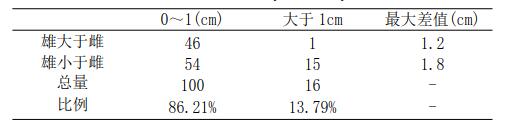
将交配的 116 对克氏原螯虾雌雄个体以体长进行比较,有 35 对雄虾体长大于雌虾,占总数的 30.17%,雄虾和雌虾体长相等的 12 对,占总数的 10.35%,雄虾体长小于雌虾的 69 对,占总数的 59.48%。雌雄虾体长差值在 0-1cm 间的虾比例很大,占到总数的86.21%(P<0.01),大于 1cm 的仅占 13.79%,最大差值为 1.8cm(表 1)。
表 1 克氏原螯虾交配对体长大小的选择
3.4 克氏原螯虾交配对体重大小的选择性
将交配的 116 对克氏原螯虾雌雄个体以体重进行比较,有 58 对雄虾体重大于雌虾,58 对雄虾体重小于雌虾,各占 50%。在交配虾中,雌雄虾的体重差值在 10g 以上的有 17对,占总数的 14.65%,最大差值为 17.68g,占总数的 28.45%,差值在 5g 以下的有 66 对,占总数的 56.90%(表 2)。
表 2 克氏原螯虾交配对体重大小的选择

3.5 克氏原螯虾雌、雄选择性
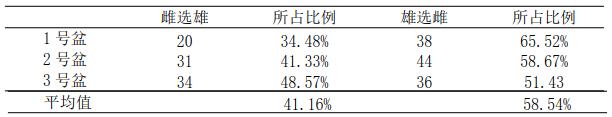
实验共观测到 203 次两性选择,其中 85 次雌选雄,占总数的 41.46%;118 次雄选雌,占总数的 58.54%。用 Origin 软件进行方差显着性分析,结果表明在 0.05 水平,没有显着性差异。表明克氏原螯虾两性选择并不是确定的雄选雌或雌选雄。
表 3 克氏原螯虾雌、雄选择
4.讨论
4.1 克氏原螯虾对交配对象地域的选择
实验中异地交配的克氏原螯虾达到了 80.17%(P<0.01),表明克氏原螯虾更趋向于与外地的虾进行交配。在没有外地虾存在或者本地虾太多的情况下会出现同地交配的现象。这也是证明克氏原螯虾在我国已经繁衍生息了 80 多年仍能保持较高的生物多样性的有力依据。不同地域的交配可以增加各地克氏原螯虾基因的交流,避免近亲繁殖,以提高自身的生物多样性,提高生长速度,增强生育能力和生存能力。
实验中 5 个地方的克氏原螯虾之间都有交配,说明克氏原螯虾之间没有生殖隔离,不同地区的克氏原螯虾属于同一个物种,可以进行繁殖育种。但是在不同地域的克氏原螯虾交配的比例不一样。各地区虾对不同地区虾的选择是随机的。
文亚峰等(2010)认为植物遗传的多样性受多种因素的影响,能确定和改变植物种群遗传多样性的内部因素包括物种的繁育系统(生殖方氏)、遗传漂变、自然选择、基因突变和基因流,同时还包括由于环境变化和人为干扰引起的种群隔离、生境片断化等外部因素。内部因素可直接作用于基因组,引起等位基因数目与频率的变化,外部因素不会直接改变基因(等位基因)数目与频率,只能通过某种间接方氏使植物群体的遗传多样性水平和遗传结构发生变化。而克氏原螯虾主要是通过自身因素,即异地交配来促进基因交流与平衡选择,从而使其遗传多样性提高。各地区克氏原螯虾没有生殖隔离在一定程度上是其可以顺利进行异地交配的前提。
4.2 克氏原螯虾对交配对象体长大小的选择
对实验中交配的 116 对克氏原螯虾雌雄个体以体长进行比较,有 35 对雄虾体长大于雌虾,占总数的 30.17%,雄虾和雌虾体长相等的有 12 对,占总数的 10.35%,雄虾体长小于雌虾的 69 对,占总数的 59.48%。雌雄虾体长差值在 0-1cm 间的虾比例很大,占到总数的 86.21%(P<0.01),大于 1cm 的仅占 13.79%,最大差值为 1.8cm。结果表明:克氏原螯虾在交配过程中,雄虾趋向于选择体长大于自身的雌虾,并且雌雄虾更倾向于体长差值较小的虾。
董朝莉(2006)在福寿螺配偶个体大小选择性初步观察中指出福寿螺与多数低等动物一样。其婚配属于乱交制,无固定配偶;雌螺对与其交配的雄螺个体大小没有选择性,而雄螺对雌螺的个体大小有选择性,倾向于与较大个的雌螺交配。雌螺个体大,总怀卵量大,卵粒直径也大,有利于受精卵的孵化和幼螺的成活,有利于其种群的繁衍和扩散。
而雌螺对雄螺没有选择性,任何一只有生殖活力的雄螺的精子数量都足以使一只雌螺的卵受精,因此雌螺不需要对雄螺个体大小进行选择。克氏原螯虾对交配对象体长大小的选择性在一定程度上与福寿螺相似,即雄虾趋向于选择体长大于自身的雌虾,而雌虾对与其交配的雄虾大小并无显着选择性。对于这点,可以类似地进行解释。即雌虾个体越大,总怀卵量越大,卵粒直径也大,有利于受精卵的孵化和幼虾的成活,有利于其种群的繁衍和扩散。而雌虾对雄虾没有选择性,任何一只有生殖活力的雄虾的精子数量都足以使一只雌虾的卵受精,因此雌虾不需要对雄虾个体大小进行选择。
而其他一些研究表明雌虾也会对雄虾进行选择。首先,雌性可能会选择大型雄性因为它们提供了重要资源,如高质量的洞穴。事实上,在生殖期间,有交配期的虾对已发现分享同一洞穴(Huner,1992),以便积极防御入侵(Figler&Peeke,2005)。大的雄性更容易赢得战斗或维护洞穴,这可能是雌性选择对较大的配偶的原因。第二个直接的好处是,雌性与大的雄性交配也许是它们能生产出更多的精子,因此,雌性可以选择它们来减少卵不受精的风险(Williams,1992)。但在最近的研究与此观点发生冲突,显示出射精量在这一物种与雄性个体的大小成反比(Rubolini 等,2006)总的来说,在克氏原螯虾对交配对象体长大小的选择上,可以确定的是雄虾偏好于体型大的雌虾,而雌虾对雄虾个体大小究竟有无选择性还有待定论。就本实验而言,雌虾对雄虾并无明显选择性。
4.3 克氏原螯虾对交配对象体重大小的选择
实验中共观测到 116 对克氏原螯虾进行交配,其中体重差值在 10g 以上的有 17 对,占总数的 14.65%,差值在 5~10g 的有 33 对,占总数的 28.45%,体型相差 5g 以内的占总数的 56.90%,数据显示体重差值越小的,交配的虾数量所占比例越高,即表明克氏原螯虾在交配时,更趋向于与自身体重比较接近的虾进行交配。
对于以上实验现象,可以做如下解释:
邱高峰等(1995)在实验中表明,克氏原螯虾的交配行为过程可分为交配前期和交配期。在交配前期,雄虾表现出特有的交配前期行为;交配期包括钳夹、翻转、横跨、交尾和分离五个阶段。雌虾被雄虾钳住后表现顺从行为。这一点说明如果雄虾与一只体重比其重太多的雌虾进行交配,就会大大增加其交配难度,交配期的各种动作进行的比较缓慢,从而无法保证其交配效率,甚至无法进行交配。张小谷等(2010)在克氏原螯虾形态性状与体重的关系中的研究表明,雌雄虾体重与体长的比值都随体长增长而逐渐增大,雄虾的体重增长相对体长增长比雌虾快。其研究说明雄性克氏原螯虾体重的增长与体长并不像雌虾那么明显,其主要一点原因就是雄虾拥有大螯,螯的重量占其体重的比例较大。大螯不仅保证了交配的顺利进行,而且在雄虾争夺配偶、领地保护、社会地位等方面具有重要意义。
本实验对克氏原螯虾的雌、雄选择性也做出了研究。结果不能说明是明显的雄选雌,只能说明雄选雌的比例大于雌选雄。L. Aquiloni & F. Gherardi(2007)研究表明体型大的虾,无论雌、雄都会被优先选择,而雄性会优先选择未交配的雌虾。本实验结果,41.46%的雌选雄及 58.54%雄选雌在一定程度上印证了其结论。雌、雄会进行相互选择,故其选择比为 1:1,而雄虾选择多出了优先选择未交配雌虾这一条件,可能会使选择比稍稍偏向于雄性。
5.参考文献
[1] 王卫民 .软壳克氏原螯虾在我国开发利用的前景 .水生生物学报[J] ,1999 ,23(4) :375-381.
[2] 王长忠 ,李 忠 ,梁宏伟 ,等 .长江下游地区4个克氏原螯虾群体的遗传多样性分析[J] .生物多样性 ,2009 ,17(5) :518-523.
[3] 刘炜 .克氏原螯虾肌肉肌苷酸含量及遗传多样性研究[D] .扬州 :扬州大学 .2008 .
[4] 周波 .性选择中的奔离过程与障碍原理[J] .遗传 ,1995 ,17(4) :43-48.
[5] Smith C ,Barber I ,Wootton R J ,et al .A receiverbias in the origin of three-spined stickleback matechoice[J] .ProcR SocLond B ,2004 ,271 :949-955.
[6] Houde A E .Sex .color andmate choice in guppies[M] .Princeton:PrincetonUniversity Press ,1997.
[7] Mc Cartney M A ,Lessios H A .Adaptive evolution of sperm bindin tracks egg incom-patibility in Neotropical Sea Urchins of the genus echinometr .MolBiol Evol ,2004 ,21(4) : 732-745.
[8] Moy G W ,Springer S A ,Adams S L ,etal .Extra-ordinary intraspecific diversity inoyster sperm bindin[J] .Proc Natl Acad Sci USA ,2008 ,105(6) :1993-1998 .
[9] 徐建荣 .福寿螺繁殖生态学与遗传多样性研究[D] .青岛 :中国海洋大学 ,2009 .
[10] 董朝莉 .福寿螺配偶个体大小选择性初步观察[J] .广西植保 ,2006 ,19(1) :4-6.
[11] 长有德 ,康乐 .昆虫在多次交配与精子竞争格局中的雌雄对策[J] .昆虫学报 2002 ,45(6):833-839.
[12] BOOMSMA J J .Split sex ratios and queen-male conflict over sperm allocation[J]
.Proc Soc Lond ,1996(263) :697-704.
[13] DARWIN C .The Origin of Species by Means of Natural Selection[M] .London :CABInternation ,1872 :1993-2002.
[14] BIRKHEAD T R ,HUNTER F M .Mechanism of sperm competition[J] ,Trends Ecol Evo,1990 ,5 :48-52.
[15] ALVAROL B ,MICHAEL D ,GREEN F .Sexual selction in insect choruses :influences ofcall power and relative timing[J] .J of Insect Behav ,2005(18) :79-83.
[16] BROWN W D ,WIDENAN J ,ANDRADAE M L B .Female choice for an indicator of malesize in the song of black horned cricket ,Oecanthus nigricornis(Orthoptera:Gryllidae:Oecanthinae)[J] .Evol ,1998 ,50(6) :2400-2411.
[17] 初世伟 ,王林波 ,姜绍赞 .红螯螯虾自然温度下人工繁育技术 ,齐鲁渔业 ,2003 ,20(5):7~8.
[18] 丛宁 ,减素绢 ,姜增华,成强,成劲松 .红螫螯虾生物学特性的观察及与克氏螯虾的比较[J] .河北水产 ,1998 ,2 :3~6.
[19] 李胜 ,赵维信 .克氏原螯虾大颚器在卵巢发育周期中的组织结构变化[J] ,上海水产大学学报 ,1999 ,8(1) :12~18.
[20] 吕佳 ,宋胜磊 ,唐建清 ,葛家春 ,潘建林 .克氏原螯虾受精卵发育的温度因子数学模型分析[J] .南京大学学报(自然科学) ,2004 ,40(2) :226~231.
[21] 罗字良 ,吴志新 ,陈孝煊 ,申学华 .红螯螫虾精巢发育的组织学研究[J] .华中农业大学学报 ,1999 ,18(1) :78~79.
[22] 文亚峰 ,韩文军 ,吴顺植 .物遗传多样性及其影响因素[J] ,中南林业科技大学学报 ,2010,30(12) :81~86.
[23] 张小谷 ,王建民 ,曹烈 ,杨建远 ,张吉勇 ,付纹琪 .克氏原螯虾形态性状与体重的关系[J] ,南昌大学学报 ,2010 ,34(2) :195~199.
[24] 邱高峰 ,诸南山 ,赖伟 .日本沼虾雄性生殖系统的研究Ⅲ :输精管内精荚的结构与形成[J] ,动物学报 ,1997 ,43(1) :68~71.
6.致谢
本论文是在导师×××教授的悉心指导下完成的。他那充满睿智的思想,开拓性的思维和渊博的知识,严谨的学风和治学态度,对科学的执着追求和忘我的工作精神以及对学生们的关心和爱护将使我受益终生。在此,谨向导师致以最诚挚的感谢和深深的敬意。
我深知科学研究是一项集体合作的工作,我能够顺利完成本科论文,有赖于许多老师和同学的支持与帮助。在此,感谢生物与食品工程学院×××老师及××师兄在我学习和工作中给予的指导和帮助。
对所有曾经在工作和学习过程中给我提供过帮助的老师和同学表示最真诚的感谢!





