
鱼类的肌间骨(Intermuscular bone)是由肌膈结缔组织连续同源骨化而来的膜骨, 通过结缔组织连在其他骨骼上, 起着加强肌节间联系的作用。我国淡水主要养殖鱼类, 如鲫鱼(Carassius carassius)、草鱼(Ctenopharyngodon idella)、鲢(Hypophthalmichthysmolitrix)等鲤科鱼类普遍存在复杂的肌间骨, 对鱼肉的食用带来极大不便。鱼类的肌间骨根据附着的部位可以分为三类, 包括髓弓小骨、椎体小骨和脉弓小骨。低等真骨鱼类普遍存在肌间骨, 且髓弓小骨形态的演化与低等真骨鱼类的系统演化进程有着一致性。国内外对鱼类肌间骨的研究多处于形态方面, 且主要集中在对不同鱼类成体的肌间骨研究, 缺乏对其发生发育的观察, 仅秉志、Bird 和 Mabee、柯中和等和吕耀平等分别对鲤科鱼类的鲤(Cyprinus carpio)、斑马鱼(Danio re-rio)、鲢、唇鱼骨(Hemibarbus labeo)肌间骨的骨化时间及发育进行了描述。
团 头 鲂 隶 属 于 鲤 形 目 (Cypriniformes), 鲤 科(Cyprinidae), 鲌 亚 科 (Culterinae), 鲂 属 (Megalo-brama), 是我国特有的重要草食性经济鱼类之一,近几年对其基因与分子标记资源的开发有了较多研究。在骨骼研究方面, 苏锦祥等对团头鲂骨骼系统的发育进行了报道, 董在杰等在对几种常见鲤科鱼类肌间骨进行分析比较的时候, 已简要概括了团头鲂成鱼肌间骨的分布和形态特征。本研究在其基础上, 通过对团头鲂仔稚鱼肌间骨骨化发育的观察, 以及对不同大小个体团头鲂肌间骨数量与形态的分析, 初步阐明了团头鲂肌间骨发育的基本规律; 同时探讨了肌间骨骨化、发育、长度与个体年龄和生长的关系。本研究结果为今后揭示团头鲂肌间骨发生与发育的分子机制提供了必要的基础资料。
1 材料与方法
1.1 实验材料
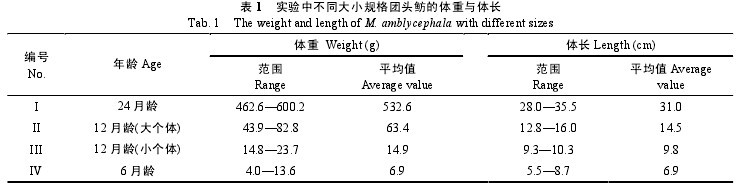
用于肌间骨染色的团头鲂仔、稚鱼样本从孵出后开始采集, 每隔 5 天取一次样本, 每次 10 尾。用95%酒精固定, 一直取到孵出后 50d, 即肌间小骨全部出现为止。形态解剖中不同大小个体的团头鲂共29 尾, 体重为(4.0—600.2) g, 体长为(5.5—35.5) cm。
按照月龄与大小将 29 尾团头鲂分为 4 组(表 1)。用于 X 光透射的成体团头鲂两尾, 体重(400—450) g,体长为(25.5—26.3) cm。以上实验鱼均取自华中农业大学水产学院团凤百容养殖基地。【表1】
1.2 仔、稚鱼骨骼染色方法
按照柯中和等的研究方法, 其中步骤(3)、(4)做以下调整: (3) 转移标本至1% KOH溶液中浸泡30min, 使组织透明, 体长大于20 mm的稚鱼则适当延长浸泡时间直至组织透明; (4) 经透明的标本转移至新的1% KOH 溶液中, 滴加茜素红染色液(1 g茜素红溶于100 mL 1% KOH 溶液中配制成染色液),直至溶液颜色变为深紫色, 染色(2—6)h (具体时间根据鱼体大小和季节温度调整), 倒掉染色液, 用1% KOH 溶液洗去组织中残余的染色液。其他步骤不变。骨骼染色的标本经体视解剖镜和高倍显微镜观察和拍照, 记录肌间骨数目、位置及形态。
1.3 肌间骨数目、形态的观察与测定
在活鱼麻醉后, 经纱布擦干鱼体后进行体重与体长的测量。测量完后用纱布将整个鱼体包好, 并用细线扎紧, 置沸水锅中煮(5—10)min。取出, 冷却后去除纱布, 剥去皮肤后, 用镊子小心剔除肌肉,依次从前往后取出肌间骨, 然后用数码相机拍照。
参考董在杰等的方法将鱼体划分为躯干部轴上肌、躯干部轴下肌、尾部轴上肌和尾部轴下肌 4 个部位, 记录不同部位肌间骨数目及形态(图 1)。用游标卡尺测定各肌间骨的长度。
1.4 X 光透射法
将研究的团头鲂送至华中农业大学校医院放射科, 进行 X 光透射拍照。观察并记录肌间骨的数量、形态与分布。
2 结果
2.1 团头鲂肌间骨的发育和骨化
在对所采样本进行肌间骨染色观察后发现: 孵出后 20 天之前的团头鲂仔稚鱼没有肌间骨存在; 第20 天的样本中, 仔稚鱼其他骨骼包括主轴骨骼及附肢骨骼已骨化完全, 游泳摄食能力完善(图版Ⅰ-A),且生长发育快的、体长为 1.33 cm 的团头鲂已出现肌间骨, 包括两根“1”形髓弓小骨与一根“1”形脉弓小骨(图版Ⅰ-B)。之后, 肌间骨不断向头部方向分化出现; 第25天, 体长为1.63 cm时, 髓弓小骨在背鳍下方出现, 形态都为“1”形(图版Ⅰ-C); 第 27 天, 髓弓小骨在背鳍前方开始出现, 而生长发育快的大个体(体长为 2.05 cm)已出现至头后方, 并在躯干轴上部分出现了少量“卜”形与“y”形肌间骨(图版Ⅰ-D、图版Ⅰ-E); 第 30 天, 体长为 1.80 cm时, 团头鲂的肌间骨已明显骨化, 形态清晰, 躯干轴上部分出现大量“y”形肌间骨(图版Ⅰ-F), 尾部出现完整明显的“1”形肌间骨(图版Ⅰ-G); 在第 40 天, 体长为 2.36 cm 时,肌间骨基本全部出现。团头鲂的髓弓小骨与脉弓小骨都是从尾部向前端依次出现, 其形态随其生长发育由尾部向头部方向逐渐复杂化。
2.2 不同大小团头鲂的肌间骨数目与长度
通过比较了 29 尾不同大小个体团头鲂肌间骨数目发现: 不同大小规格的 4 个组合团头鲂肌间骨数目并无显着差异(P>0.05), 团头鲂肌间骨数目在108—129, 左右两侧肌间骨数目在 53—67。实验中发现每尾鱼左右两侧的肌间骨数目不完全相等, 但总体上左右两侧肌间骨的数目接近(表 2)。
团头鲂仔稚鱼阶段其肌间骨长度是随其生长明显增加的, 本研究对不同月龄团头鲂肌间骨长度进行了解剖测量, 结果显示肌间骨长度仍然随其生长显着增加(P<0.05)(图 2)。团头鲂躯干轴上肌中的肌间骨显着长于尾部肌肉中的肌间骨(P<0.05), 躯干轴下肌中的肌间骨最短, 尾部轴上肌与尾部轴下肌中的肌间骨长度无明显差异。12 月龄小个体团头鲂中躯干上肌间骨平均长度为(7.83±1.16) cm, 尾部上肌间骨长度为(7.14±1.09) cm, 尾部下肌间骨长度为(7.02±0.77) cm, 躯干下肌间骨长度为(5.96±0.23) cm。而 12 月龄的大个体团头鲂肌间骨要显着长于小个体团头鲂(P<0.05), 其中躯干上肌间骨平 均 长 度 为(11.70±2.36) cm, 尾 部 上为(10.92±1.47) cm, 尾部下为(10.61±1.81) cm, 躯干下为(9.65±0.12) cm。【表2.图2】


2.3 团头鲂肌间骨形态与分布
本研究结果将团头鲂肌间骨形态分为“1”形、“卜”形、“y”形、一端多叉形、两端多叉形和“(”形 6种类型。在团头鲂仔稚鱼的连续染色观察中发现:
“卜”形表现为“y”形、一端多叉形和两端多叉形发育过程中的过渡形态。团头鲂左右两侧不同部位的肌间骨数目有差异, 躯干部轴上肌中的肌间骨数目最多, 平均为 22 根, 且随着团头鲂个体的变大, 在躯干部轴上肌的分叉肌间骨中发现了两端分叉与一端分为 3—4 枝的情况, 肌间骨形态多样性增加(表 3)。
尾部轴上肌与轴下肌中的肌间骨数目非常相近, 且多为“1”形。躯干部轴下肌中的肌间骨最少, 且均分布在腹腔与尾部的过渡区域。肌间骨整体形态以“1”形和“y”形为主(表 3); 在此基础上, 出现 1—4 个分支, 其中一端多叉形、两端多叉形的肌间骨多存在于12月龄大个体与24龄成鱼, 且形态最多样(图3)。
X 光透射图像清晰地展现了团头鲂肌间骨在鱼体中的发生区域、与主轴骨骼及附肢骨骼的相对分布位置、数量及形态(图版Ⅱ)。
3 讨论
3.1 团头鲂肌间骨的骨化与发育
鱼类骨骼的骨化是一个非常复杂的过程。在对鱼类骨骼系统的研究中, 多为椎骨的研究, Mabee等对鱼类椎骨发生及椎骨与鱼类进化之间的关系进行了研究, Bird 和 Mabee对斑马鱼的椎骨的发育演化进行了研究。国内对鱼类肌间骨的研究多处于形态方面的研究, 且处于初步研究阶段, 对于肌间骨的形成原因、演化规律、发育的分子机制等方面的研究尚未开展。肌间骨形态发生规律的研究, 是研究其发育分子机制的基础。柯中和等研究发现鲢的肌间骨骨化的时间较晚, 在主轴骨骼及附肢骨骼出现之后才开始骨化, 其第一枚肌间骨在 43dpf (体长为 10.50 mm)时出现。吕耀平等研究发现唇鱼骨的肌间骨在 35dpf (体长为 23.67 mm)开始在尾部区域骨化。本研究发现团头鲂在孵出后 20d 肌间骨骨化出现时, 其主轴骨骼及附肢骨骼已骨化完全,鱼体运动觅食能力已基本完善, 与前者研究相一致,且同一时期肌间骨出现与尚未出现的鱼运动摄食能力并无显着差异。因此, 在肌间骨形成之前抑制其骨化出现, 理论上不会对鱼体主要骨骼的分化发育与生长产生影响; 抑制团头鲂肌间骨骨化, 培育无肌间骨的团头鲂, 在理论上应该是可行的。
从团头鲂仔稚鱼肌间骨染色观察实验中发现团头鲂的脉弓小骨与髓弓小骨都是从“1”形肌间骨逐步分化发育而来的, 形态复杂的髓弓小骨按照“1”形→“y”形→多叉形的方向分化发育。幼鱼在孵出后20d 左右(体长约 1.33 cm)出现第一根肌间骨; 40d 左右(体长约 2.1 cm), 肌间骨基本完全出现; 之后随着鱼体的生长, 躯干轴上肌的髓弓小骨形态不断分化复杂, 直到成体团头鲂的生长过程中, 肌间骨形态依然在分化。这说明肌间骨的分化是伴随鱼体生长不断进行的, 形态越复杂的肌间骨形成越晚。此外,同一时期样本仔稚鱼所含的肌间骨数量与分化程度不完全相同, 第 25 天、体长 1.45 cm 的仔稚鱼已经有超过 15 根的肌间骨, 而 1.36 cm 的仔稚鱼尚未出现肌间骨; 第 27 天、体长 2.05 cm 的仔稚鱼躯干轴上肌已经分化出“y”形肌间骨, 而第30 天、体长1.33 cm的仔稚鱼仅存在尾部“1”形肌间骨。这些结果表明团头鲂肌间骨出现与分化的时间受生长发育的影响大于日龄的影响。
3.2 团头鲂肌间骨的数目与长度
本研究发现不同大小规格的团头鲂(5.5—35.5 cm)肌间骨数目无显着差异(P>0.05), 在 108—129, 平均为 119 根, 与董在杰等对团头鲂等鲤科鱼类肌间骨数目的研究结果相近。同时, 团头鲂的肌间骨数目并不是呈两侧对称分布的, 但十分接近。这与目前国内学者孟庆闻等的研究是不一致的, 其在对鲢的肌间骨研究中发现肌间骨是两侧成对分布的。但后来国内学者的研究中发现鱼类的肌间骨并不呈两侧对称分布, 与本研究得到的结论是相同的。
团头鲂的肌间骨是由肌膈结缔组织连续同源骨化而来的膜骨, 肌间骨的长度与团头鲂的体重和体长呈正相关。本研究发现团头鲂躯干轴上肌肌肉最多, 肌间骨的长度也最长; 相反, 躯干轴下肌由于腹腔的存在其肌肉最少, 肌间骨的长度最短。这间接说明了团头鲂肌间骨的长度与肌肉的分布存在密切的联系, 肌肉多的躯干轴上肌间骨长度要大于肌肉少的躯干轴下肌间骨。Johnson 和 Patterson对鱼类肌间骨的研究认为: 鱼类肌间骨可能起着传导肌肉力量与支撑肌肉的作用。因此, 可以推断团头鲂躯干轴上与轴下肌中的肌间骨长度差异可能是因肌肉的分布差异造成的, 肌间骨的生长与肌肉的生长是同步的, 这与本研究中肌间骨发育与鱼体生长发育相关联的结果是互相印证的。
3.3 团头鲂肌间骨的形态与分布
鱼类肌间骨的形态表现出多样性, 且与存在部位有一定关系。本研究发现团头鲂躯干上肌中的肌间骨形态最为复杂, 而尾部肌肉中的肌间骨要相对简单, 这与董在杰等对团头鲂肌间骨的研究结果是一致的。关于肌间骨的形态类型, Patterson 和Johnson从系统进化的角度分析认为骨舌鱼总目最原始的肌间小骨形态是没有分叉的, 在此之后的骨鳔总目, 包括鲤形目的鲤科、胭脂鱼科、鳅科鱼类,脂鲤目鱼类, 鲇形目鱼类等的肌间骨都是从“1”形肌间骨演化而来的。吕耀平等通过观察各种低等真骨鱼类肌间小骨的形态, 认为肌间骨的演化是从简单的“1”形逐步演化为复杂的“卜”形、“y”形、两端多叉形和树枝形。本研究根据对不同个体大小团头鲂肌间骨形态的观察, 认为团头鲂肌间骨的演化过程与吕耀平等报道的研究结果是一致的。在本研究中还发现团头鲂肌间骨形态存在“(”形, 根据其所处位置及形态特征, 推测其为受到肌肉张力及发育限制后的“卜”形的未发生体或者“1”形的变形体。
参考文献:
[1] Xie C X. Ichthyology [M]. Beijing: China Agricultural Press.2010, 56—57 [谢从新. 鱼类学. 北京: 中国农业出版社.2010, 56—57]
[2] Ma L R, Dong Z J, Su S Y, et al. The research progress onintermuscular bones of teleosts [J]. Jiangsu AgriculturalScience, 2012, 40(4): 234—235 [马良骁, 董在杰, 苏胜彦,等. 鱼类肌间刺的研究进展. 江苏农业科学, 2012, 40(4):234—235]
[3] Meng Q W, Su Q W, Li W D. Comparative Anatomy ofFishes [M]. Beijing: Science Press. 1987, 102—103 [孟庆闻,苏锦祥, 李婉端. 鱼类比较解剖. 北京: 科学出版社. 1987,102—103]
[4] Patterson C, Johnson G D. The Intermuscular Bones andLigaments of Teleostean Fishes [M]. Smithsonian Contri-bution to Zoology. 1995, 1—85
[5] Lü Y P, Bao B L, Jiang Y, et al. Comparative analysis ofintermuscular bones in lower teleosts [J]. Journal ofFisheries of China, 2007, 31(5): 661—668 [吕耀平, 鲍宝龙,蒋燕, 等. 低等真骨鱼类肌间骨的比较分析. 水产学报,2007, 31(5): 661—668]





