

在放入含1 M的DMSO保存液中之前,以每次0.25M浓度逐级增加至1M,每增加一级平衡孵育5min,在最终浓度内平衡10min.采用同样的步骤进行DMSO的稀释。在放入DMSO之前或之后,在无DMSO的鱼Ringee液中平衡5min.
我们对完整的卵细胞进行了过冷点与冰点的测定,采用铜一康铜温差电偶记录过冷点和冰点的温度,指示部分采用国产AC15/2型直流复射式检流计,配备国产ZX21型旋转式电阻箱。
降温系采用自制简易降温仪(图1),致冷剂为碎冰和食盐的混合物,其原理是通过致冷剂冷却烧杯内的酒精,从而使样品致冷。由半导体温度计测量保存液的温度。在进行冷冻前与冷冻后,均以每5min增加或减少0.25M的步骤平衡受精卵(室温),而后放到简易降温仪内降温,降温曲线如图2所示。冷冻所达的温度分别为一6℃,一10℃,一15℃,并在其最低温度内保存10min.冷冻结束后,以10~20℃/min的复温速度解冻鱼卵(8℃),经过DMSO平衡之后移入水中激活,2小时后计数其直接成活率,孵化至发眼期,计数其发眼卵数。


冷冻卵活力的签定标准为:卵对水的活性或变硬处理有反应,则认为卵是活的;变得透明,不发生机械变形和产生卵周隙的卵,被认为是变硬的卵。
结果:
1.冷冻对受精卵孵化效果的影响:
在未冻结的保存液内,卵的外形直观上无变化,但当保存液有冰晶产生时,有些卵变白,解冻时,这些卵又变得清晰,但移到水中激活后,这些卵又重新变白。没有变化的卵,经水激活后形态正常。经冷冻保存之后的卵,透明度增高,卵细胞内,油滴多聚集在动物极之侧。
冷冻实验由于系分批进行,故全部实验数据均与相应的对照值相关。
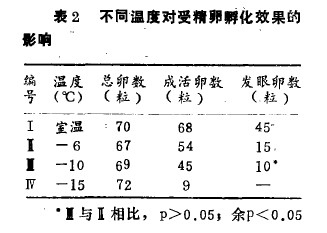
冷冻组的样品从室温降至一5℃这一阶段,含有冷冻保护剂的保存液并未冻结,在保存液内形成冰晶时,温度上升至一4℃左右,其所持续的时间在1min之内。样品的中心重新冷至所需温度是以一0.2℃/min的速度进行的,并在最低温度保存l0 min,结果见表2.
2.过冷点与冰点的测定:本试验所测得的过冷点是不恒定的,其值均在 -7.5℃以上;但其冰点值均在-1.7 ~ -1.8℃之间(表3)。

讨论此次实验全部采用没被水激活的受精卵,据鱿鱼(Salmo Salar)的研究,就水透过卵黄外周膜而言,未激活比激活的卵渗透系数高,由此可知,冷冻期间未激活的卵可能脱水较好,因而胞内冻结而引起损害的几率最小。此外精子与卵子的细胞质接触后,由于未激活的卵受精过程被阻滞,所以这些卵子具有统一的发育期。
Zell、Erdahl和Grahum也曾用受精、但未激活的卵作冷冻研究。受精卵在体腔液或鱼Ringer液中保留有受精性,尽管在鱼Ringer液中能够受精,但只有将其移到新鲜水中时它才有活性和变硬。渗透压对受精卵的影响,可采用的资料有限,但从B、Harvey(1983)和J、Stoss(1983)的资料来看,其维持在240一450mosm可耐受24小时。本次试验我们所采用的鱼Ringer液的渗透压维持在250mosm左右。
冷冻前和冷冻后添加DMSO的主要目的是防止渗透冲击,使抗冻剂充分渗透。
抗冻剂的穿透性与该物质的分子量是成反比的。J、Stoss(1983)证明:冷冻前DMS0在2 M和4 M对银大马哈鱼卵有害,几乎不能存活,但含2 M的DMSO样品被冷冻至一11.8℃时,仍可观察到部分卵活到发眼期。逐步增加DMSO的浓度,可以减少或消除不利影响。冷冻保护剂可使细胞脱水时间更长,延缓细胞内冰的形成。现普遍认为IM的DMSO效果似乎较好,故此,我们所采用的DMSO浓度亦为1 M。
Harvey和Smith(1982)测得硬头缚卵的内容物和完整卵的冰点近似,其值均为一1.7℃,这与我们的结果相比是一致的。很多类型的细胞质在一1℃便结冻。蛙鱼卵的冰点较低,无异是由它们卵黄内容物高的缘故,过冷点值的不恒定,是与降温速度有关,降温速度越快,过冷状态越不稳定,过冷点越小,反之也是如此。
冷冻和解冻期间,存活率减少的主要原因是细胞内冰形成,而冷冻程序主要在于充分地脱去细胞内的水分,使细胞内冰形成的几率减至最小。
稀释液与卵细胞自然冻结的温度为一4℃,这与Harvey和Smith(1982)所得的结果是相近的。在一6℃DMSO有效地保护了结冰过程中的受精卵,然而进一步冷却至一15℃多数细胞内结冰,这说明所用的冷却速度使细胞脱水不充分,Mazur指出:适宜的冷却速度和细胞的直径大致是相关的。故此,我们所用的降温速度可能是不适合的。Zell(1978)在没有抗冻剂的情况下,以5℃/min的冷却速度将溪红点蛙受精卵降至一5℃,孵化率为20%,在含有抗冻剂(8%DMSO,10%PVP)的情况下,在相同条件下获得70%的存活率,Erdahl和Graham(1980)以5℃/min降温速度,在一20℃保存,持续时间不清,含14%DMSO或14%甘油的保存液中在16粒欧傅鱼卵中获得69%的发眼率,但在更低的温度下,没有获得成功。由于Zell(1978)每次试验使用的卵量较多(75~750),卵置于最低温度不到5 min,这样,样品内可能会产生温度梯度,中心温度可能未达到最低温度,所以这些结果与我们的资料相比是困难的。在我们的研究中多数卵细胞内冻结,因此在含有DMSO的保存液结冰之后,在一10℃~一15℃之间坏死,与Stoss(1983)的结果是较相近的。Harvey和Smith(1982)的研究结果也表明,在10%的DMSO的清况下,保存硬头蹲卵在一16℃,鱼卵细胞内亦结冰。最低温度时的存活率也是一样不能保证以后的发育率,这一点在我们的资料中也可体现出来。
总的来讲,寻求更慢的降温速度与合理的改变解冻速度及实验程序,使细胞内脱水更充分,可能使鱼卵的冷冻获得进一步的成功,有待于新的探索。
参考文献:
[1] J.Stoss and EMDonaldson(李禾译):Aquaeulture1983,31(1):51~65
[2] Brian Harvey and MJ Ashwood Smith: Cryobiology 1982,19(1):29~40
[3] Brian Harvey ,et al: Can Teeh Rep Fish Aquat Sei 1983, 1222:1~9
[4] 李楚杰:冷伤人民卫生出版社。1980,48一50
[5] Ginsburg AS: J.Embryol ExP MorPhol 1963, 11:13~33
[6] Zell SR:Ann Biol Anim Bioehim BiaPhys 1978, 18:1089~1099
[7] Erdahl, DA, and Graham, EF: In “Proeeedings International Conferenee on Animal ReProduetion and Artifiecal Insemination, 1980; 317~326
[8] Mazur, P: Cryobiology 1977,14:251~272
[9] Mazur, P: Cryobiology Seieoee 1970, 168:939~949





