摘要:梅花鹿Cervus nippon为国家Ⅰ级重点保护动物,四川铁布梅花鹿自然保护区分布有我国现存最大的梅花鹿野生种群。2011年6-9月,采用样方法对保护区梅花鹿夏季栖息地选择进行了调查。共布设109个样方(利用样方61个,对照样方48个),测量并比较了海拔、坡度等20个生境因子。结果显示,梅花鹿偏向利用的植被类型为灌丛、草甸、针叶林,同时选择隐蔽度较高的生境;此外,距水源距离、灌木高度、灌木盖度、灌木密度、草本高度和草本盖度6个连续变量在利用样方和对照样方之间差异有统计学意义。回归模型分析结果表明,梅花鹿夏季偏好选择草本盖度大、距离水源和林缘较近的生境,拒绝远离水源和林缘的生境。
关键词:梅花鹿; 栖息地选择; 铁布梅花鹿自然保护区;
作者简介: 赵成(1986-),博士,讲师,生态学专业,主要从事动物生态保护研究,E-mail:zhaochengbox@163.com; 胡杰, 博士, 教授, 主要从事珍稀野生动物保护遗传和生态学研究, E-mail:hu_jiebox@163.com;
Summer Habitat Selection of Cervus nippon in the Sichuan Tiebu Sika Deer Nature Reserve
Abstract:Sika deer (Cervus nippon) is classified as a class Ⅰ key protected wild animal species in China. The largest wild population of C. nippon is distributed in the Sichuan Tiebu Sike Deer Nature Reserve. From June to September 2011, quadrat sampling method was used to investigate the summer habitat selection of C. nippon in the reserve. Twenty habitat variables, such as elevation and slope gradient, were assessed in 109 quadrats. Of the 109 qudrats, 61 sites were used for the survey and 48 were set as control. The results showed that C. nippon preferred bush, meadow, and coniferous forest with high concealing condition. There were significant differences between the used and control quadrats in 6 continuous variables, such as the distance to water, shrub height, shrub cover, shrub density, herb height, and herb cover. The result of logistic regression analysis showed that C. nippon prefers the habitat with higher herb cover, shorter distance to water and forest edge in summer.
Keyword:Cervus nippon; habitat selection; Tiebu Sika Deer Nature Reserve;
梅花鹿Cervus nippon隶属于偶蹄目Atiodactyla鹿科Cervidae,国家Ⅰ级重点保护野生动物,历史上几乎遍布我国华东、华北、华中、华南、西南和青藏高原的东部(Whitehead,1993;郭延蜀,郑慧珍,2000),由于气候环境的改变以及人为影响,目前中国仅存3个梅花鹿亚种:东北亚种C. n. hortulorm、华南亚种C. n. kopschi和四川亚种C. n. sichuanicus,分布于黑龙江、吉林、辽宁、安徽、浙江、江西、四川和甘肃迭部交界区域(郭延蜀,2000;宋延龄,刘志涛,2005);其中,四川铁布梅花鹿自然保护区分布有我国现存最大的野生种群(戚文华等,2014)。
栖息地是野生动物赖以生存的场所,了解和掌握濒危物种栖息地的利用特征是制定保护和管理策略的基础(孟秀祥等,2011;Zhang et al.,2011;Khadka & James,2016)。迄今为止,国外学者对梅花鹿栖息地利用方面的研究成果较为丰富(Asada & Ochiai,1996;Borkowski,2000;Yokoyama et al.,2000;Sakuragi et al.,2003;Diaz et al.,2017;Endo et al.,2017);国内关于梅花鹿栖息地利用方面的研究相对较少,且主要集中于华南亚种(付义强等,2006;李佳等,2015;Li et al.,2017),针对四川亚种栖息地利用方面的研究则更少(Zhao et al.,2014)。
由于环境的差异,不同地理种群的梅花鹿所面临的生存压力也不一样,进而导致影响其栖息地选择的主要因素也存在一定差异。例如,生活于俄罗斯西部的梅花鹿主要选择的生境为蒙古栎Quercus mongolica林、落叶阔叶林和红松Pinus koraiensis针阔混交林(Aramilev,2009);分布于吉林珲春东北虎国家级自然保护区的东北亚种却不偏好红松针阔混交林和蒙古栎林(杨海涛等,2018);而分布于江西的华南亚种不偏好森林,而偏好灌草丛(付义强等,2006;Li et al.,2017)。鉴于此,作者对四川铁布梅花鹿自然保护区梅花鹿夏季的栖息地选择进行了研究,以期进一步丰富该亚种的生态生物学资料,揭示影响其生境选择的关键因素,并为其保护与管理提供参考。
1、研究区域与研究方法
1.1 研究区域概况
四川省若尔盖县铁布梅花鹿自然保护区(102°52′~103°08′E,33°58′~34°11′N)位于青藏高原东沿,总面积约260 km2.区内地貌属中切割山原,谷底海拔2 400 m,峰岭海拔3 000 m左右,最高可达3 850 m.受西风环流及东南季风的影响,夏季温凉、冬季寒冷、干湿季明显,年均降水量625.5 mm.区内植被垂直分布明显,且坡向影响强烈。在河岸及河漫滩地带以柳Salix spp.为主,其间分布有沙棘Hippophae rhamnoides、小檗Berberis spp.、高丛珍珠梅Sorbaria arborea等的河滩灌丛;阴坡以油松Pinus tabulaeformis林、粗枝云杉P. asperata林、紫果云杉P. purpurea林、岷江冷杉Abies faxoniana林、白桦Betula platyphylla林和山杨Populus davidiana林为主;阳坡是由小檗、沙棘等组成的灌丛草甸和山坡灌丛(郭延蜀,2000)。森林、灌丛、草甸构成的复合生境为梅花鹿生命活动的各个阶段提供了适宜的生存环境(Zhao et al.,2014)。
1.2 调查方法
2011年6-9月在保护区设置20条垂直于等高线的样线,每条长2 km,间距大于2 km,且尽可能覆盖区内各种植被类型。在满足以下条件时设置样方:1)样线的起始点与终点,2)发现梅花鹿实体或新鲜粪便、尸体等痕迹处,3)植被类型发生明显变化,4)若无上述情况,则海拔每上升100 m或行走1 h设置1个样方(张文广等,2007)。
样方设置采取参考Wei等(2000)的方法:1个1 m×1 m的正方形,1个20 m×20 m的正方形和2个彼此垂直的2 m×10 m的长方形,另在20 m×20 m正方形样方中每1/4小样方(10 m×10 m)的中心,分别设1个1 m×1 m的正方形样方。每个样方记录如下信息:
坐标:用GPS记录20 m×20 m的正方形样方中心的地理坐标。
样方类型:分为1和0,1表示发现梅花鹿实体(状态:觅食、卧息等)或其粪便、食痕等的样方,0表示没发现梅花鹿及其痕迹的样方(对照样方)。
坡向:东、南、西、北和无坡向。
坡位:上坡、中坡和下坡。
坡度:用坡度仪直接测量。
海拔:在大样方的中心点由GPS测量。
距水源距离:根据GPS定位,后期结合地形图和卫星影像图测量距离。
距林缘距离:记录样方距林缘的垂直距离,从林缘到林内取“+”值,从林缘到林外取“-”值。
隐蔽度:在每个大样方的对角线上,相距30 m处,各执一花杆(高1.5 m),对视对方花杆,记录所见比例,取2次之平均数。分为4级:<25%(所见杆比例>75%)、25%~50%(所见杆比例为50%~75%)、50%~75%(所见杆比例为25%~50%)、>75%(所见杆比例<25%)。
植被类型:针叶林、阔叶林、灌丛、草甸、耕地和其他。
乔木密度:大样方内乔木的数量。
乔木高度:大样方内所有乔木的平均高度。
乔木胸径:测量4个10 m×10 m正方形样方中距中心点最近的乔木的胸径,并计算其平均值。
灌木密度:2个2 m×10 m长方形样方的灌木平均数。
灌木高度:2个2 m×10 m长方形样方的灌木平均高度。
灌木盖度:2个2 m×10 m长方形样方的灌木平均盖度。
草本高度:5个1 m×1 m正方形样方的草本平均高度。
草本盖度:5个1 m×1 m正方形样方的草本平均盖度。
距居民区距离:根据GPS定位,后期结合地形图和卫星影像图测量距离。
距大路距离:指距机动车行驶道路的距离,根据GPS定位,后期结合地形图和卫星影像图测量距离。
1.3 数据分析
针对植被类型、隐蔽度、坡度、坡向4个离散型变量,采用卡方检验分析利用样方与对照样方的差异性。
通过单样本K-S检验分析数值型变量是否符合正态分布,如果符合正态分布,则使用独立样本t检验分析利用样方和对照样方的差异性,否则,使用Mann-Whitney U检验。
通过信息理论方法(Burnham & Anderson,2002)来确定预测梅花鹿栖息地选择的关键因子。为保证变量的独立性,对所有变量进行相关分析,对相关系数绝对值>0.5的变量进行筛选,保留其中更具生物学意义的变量进入下一步分析。使用Akaike信息标准校正小样本量(AICC)评估所有可能的变量组合,并通过ΔAICC值进行模型筛选;对ΔAICC<2的模型进行模型平均,然后比较每个变量的相对重要性和95%置信区间,以此确定关键的影响因子(Wang et al.,2018)。数据分析在SPSS 20和R中完成。
2、结果
2.1 利用样方和对照样方各因子的差异显着性分析
共设置梅花鹿利用样方61个,对照样方48个。
对保护区梅花鹿夏季利用样方和对照样方中4个离散型变量分析的结果表明,植被类型和隐蔽度之间的差异有统计学意义,而坡向和坡位之间的差异无统计学意义(表1)。梅花鹿偏好灌丛、草甸、针叶林和隐蔽度较高的生境。
表1 铁布梅花鹿自然保护区梅花鹿夏季利用样方和对照样方中离散型变量比较
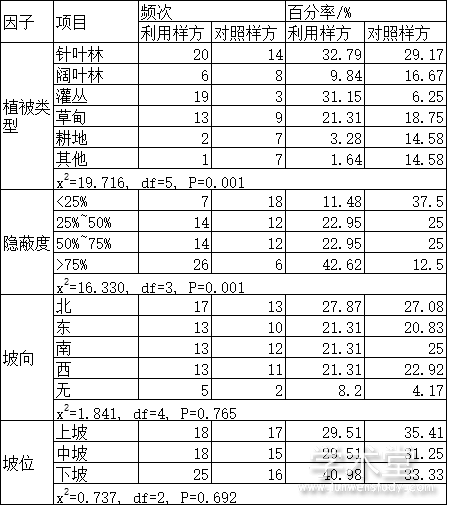
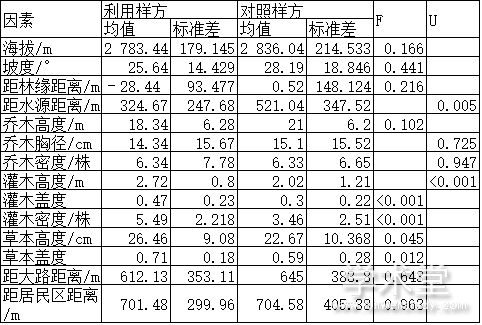
对14个连续性变量分析的结果表明,距水源距离、灌木高度、灌木盖度、灌木密度、草本高度和草本盖度6个变量在利用样方和对照样方之间的差异有统计学意义,其他变量之间的差异无统计学意义(表2)。
表2 铁布梅花鹿自然保护区梅花鹿利用样方和对照样方中连续型变量比较
2.2 逻辑斯蒂回归模型
相关分析结果发现,灌木盖度和灌木密度之间的相关系数较大(r=0.838,P<0.001)。鉴于灌木密度既能反映样方的隐蔽度又能代表样方的食物量情况,仅将灌木密度纳入模型进行分析。各回归模型中ΔAICC<2的有3个,共涉及11个变量(表3)。对ΔAICC<2的模型进行模型平均结果表明,影响梅花鹿夏季栖息地选择的主要因子为草本盖度、距林缘距离和距水源距离;且梅花鹿偏好选择草本盖度大、靠近水源和林缘的栖息地,回避远离水源、林缘的生境。
表3 最优逻辑斯蒂回归模型集合
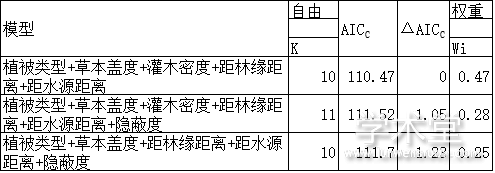
注: 植被类型为分类变量, 包含针叶林、阔叶林、灌丛、草甸、耕地和其他Note: vegetation types are categorical variables, including coniferous forests, broadleaved forests, shrubs, meadows, cultivated land, and others
表4 模型平均结果
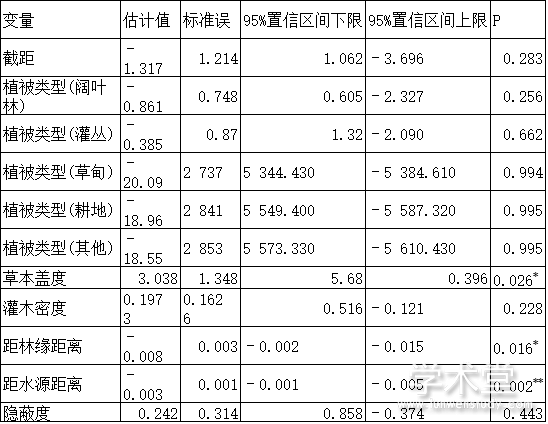
注: 植被类型(针叶林)作为固定参考因子Note: vegetation type (coniferous forest) as a fixed reference factor
3、讨论
研究表明,食物是影响动物栖息地选择的最重要因素(李伟等,2010;赵成等,2012;郑维超等,2012;王征,张旭晖,2014;刘鹏,张薇薇,2017)。郭延蜀(2001)研究发现,草甸和灌丛为梅花鹿四川亚种的主要食物基地。许多鹿科动物都会选择隐蔽性较好的生境避开天敌(刘振生等,2004;游章强等,2014)。本研究表明,梅花鹿通常偏好灌木和草本繁茂、隐蔽度较高的生境。
水源对野生动物的分布有明显的影响,鹿科动物多偏好在距水源较近的区域活动(胡杰等,2018),梅花鹿也不例外(付义强等,2006;Zhao et al.,2014)。本研究结果显示,梅花鹿偏好选择距离水源较近的生境,这进一步印证了水源对梅花鹿分布的重要性,即便是在雨水相对丰富的夏季,梅花鹿依然偏好近水生境。郭延蜀(2000)曾指出,梅花鹿四川亚种多出现在距离水源30~550 m的范围内,本研究中梅花鹿夏季利用样方距离水源为324.67 m±247.68 m,与其类似。此外,作为典型的林缘动物,梅花鹿利用的栖息地也相对靠近林缘地带。
研究表明,除了食物、隐蔽地和水源,人为干扰也是影响野生动物栖息地利用的重要因素(刘振生等,2004;冉江洪等,2006;游章强等,2014;Zhao et al.,2017;胡杰等,2018)。本研究显示,人为干扰在梅花鹿利用样方和对照样方之间的差异无统计学意义,未进入最优模型,可能有以下两方面因素:一方面,区内道路和居民点多依水而建,梅花鹿对水的需求致使其无法长期回避人为活动频繁的区域;另一方面,区内尚有大量居民从事生活生产活动,人鹿关系密切,梅花鹿已适应一定程度的人为干扰。
本研究结果表明,分布于铁布梅花鹿自然保护区的梅花鹿四川亚种在夏季偏好选择隐蔽度高、距离水源和林缘较近、灌木和草本生长繁茂的生境。鉴于此,加强管护靠近水源和林缘的灌草丛,是保护梅花鹿四川亚种栖息地的关键。
参考文献
[1] 付义强, 贾小东, 胡锦矗, 等。 2006. 江西桃红岭自然保护区夏季梅花鹿对生境的选择性[J]. 四川动物, 25(4): 863-865.
[2] 郭延蜀, 郑慧珍。 2000. 中国梅花鹿地史分布、种亚种的划分及演化历史[J]. 兽类学报, 20(3): 168-179.
[3] 郭延蜀。 2000. 四川梅花鹿的分布、数量及栖息环境的调查[J]. 兽类学报, 20(2): 81-87.
[4] 郭延蜀。 2001. 四川梅花鹿食性的研究[J]. 四川师范学院学报(自然科学版), 22(3): 112-119.
[5] 胡杰, 姚刚, 黎大勇, 等。 2018. 卧龙国家级自然保护区水鹿夏季生境选择[J]. 兽类学报, 38(3): 277-285.
[6] 李佳, 李言阔, 缪泸君, 等。 2015. 桃红岭国家级自然保护区梅花鹿和野猪秋季生境选择差异[J]. 四川动物, 34(2): 300-305.
[7] 李伟, 周伟, 刘钊, 等。 2010. 云南大中山黑颈长尾雉栖息地选择周年变化[J]. 动物学研究, 31(5): 499-508.
[8] 刘鹏, 张薇薇。 2017. 官山自然保护区白颈长尾雉季节性生境选择[J]. 生态学报, 37(18): 6005-6013.
[9] 刘振生, 曹丽荣, 翟昊, 等。 2004. 贺兰山区马鹿对冬季生境的选择性[J]. 动物学研究, 25(5): 403-409.
[10] 孟秀祥, 潘世秀, 惠岑怿, 等。 2011. 甘肃兴隆山自然保护区濒危马麝(Moschus sifanicus)秋季利用生境特征[J]. 应用与环境生物学报, 17(3): 412-417.
[11] 戚文华, 蒋雪梅, 杨承忠, 等。 2014. 四川梅花鹿繁殖行为[J]. 生态学报, 34(22): 6548-6559.
[12] 冉江洪, 曾宗永, 刘世昌, 等。 2006. 四川大相岭大熊猫种群及栖息地调查[J]. 四川大学学报(自然科学版), 43: 889-893.
[13] 宋延龄, 刘志涛。 2005. 珍稀动物梅花鹿及其研究[J]. 生物学通报, 40(7): 1-3.
[14] 王征, 张旭辉。 2014. 野生动物觅食地选择的研究进展[J]. 生态学杂志, 33(11): 3150-3156.
[15] 杨海涛, 谢冰, 韩思雨, 等。 2018. 吉林珲春自然保护区梅花鹿种群多度的季节分布及其影响因素[J]. 北京师范大学学报(自然科学版), 54(4): 498-505.
[16] 游章强, 唐中海, 杨远斌, 等。 2014. 察青松多白唇鹿国家级自然保护区白唇鹿对夏季生境的选择[J]. 兽类学报, 34(1): 46-53.
[17] 张广文, 唐中海, 齐敦武, 等。 2007. 大相岭北坡大熊猫生境适宜性评价[J]. 兽类学报, 27(2): 146-152.
[18] 赵成, 李艳红, 胡杰, 等。 2012. 嘉陵江中游长嘴剑鸻冬季觅食地选择[J]. 四川动物, 31(1): 22-26.
[19] 郑维超, 黎大勇, 谌利民, 等。 2012. 唐家河国家级自然保护区川金丝猴冬季栖息地选择[J]. 四川动物, 31(2): 208-211.
[20] Aramilev VV. 2009. Sika deer in Russia[M]. Tokyo: Springer,.
[21] Asada M, Ochiai K. 1996. Food habits of sika deer on the Boso Peninsula, central Japan[J]. Ecological Research, 11: 89-95.
[22] Borkowski J. 2000. Influence of the density of a sika deer population an activity, habitat use, and group size[J]. Canadian Journal of Zoology, 78: 1369-1374.
[23] Burnham K, Anderson D. 2002. Model selection and multimodel inference: a practical information-theoretic approach[M]. New York: Springer.
[24] Diaz A, Walls S, Whitmarsh D, et al. 2017. Habitat selection of invasive sika deer Cervus nippon living in a UK lowland heathland-woodland-grassland mosaic: implications for habitat conservation management[J]. Journal of Scientific Research, 17(3): 1-15.
[25] Endo Y, Takada H, Takatsuki S. 2017. Comparison of the food habits of the sika deer (Cervus nippon), the Japanese serow (Capricornis crispus), and the wild boar (Sus scrofa), sympatric herbivorous mammals from Mt. Asama, central Japan[J]. Manmal Study, 42: 131-140.
[26] Khadka KK, James DA. 2016. Habitat selection by endangered Himalayan musk deer (Moschus chrysogaster) and impacts of livestock grazing in Nepal Himalaya: implications for conservation[J]. Journal for Nature Conservation, 31: 38-42.
[27] Li J, Li YK, Liu WH. 2017. Autumn bed selection by sika deer (Capricornis crispus) in the Taohongling National Nature Reserve, China[J]. Russian Journal of Ecology, 48(4): 384-391.
[28] Sakuragi M, Igota H, Uno H, et al. 2003. Seasonal habitat selection of an expanding sika deer Cervus nippon population in eastern Hokkaido, Japan[J]. Wildlife Biology, 9(1): 141-153.
[29] Wang B, Xu Y, Zhang B, et al. 2018. Overlap and selection of dust-bathing sites among three sympatric montane galliform species[J]. The Auk, 135(4): 1076-1086.
[30] Wei FW, Feng ZJ, Wang ZW, et al. 2000. Habitat use and separation between the giant panda and the red panda[J]. Journal of Mammology, 81(2): 448-455.
[31] Whitehead Gk. 1993. The encyclopaedia of deer[M]. UK: Swan Hill.
[32] Yokoyama M, Kaji K, Suzuki M. 2000. Food habits of sika deer and nutritional value of sika deer diets in eastern Hokkaido, Japan[J]. Ecological Research, 15: 345-355.
[33] Zhang ZJ, Swaisgood RR, Zhang SN, et al. 2011. Old-growth forest is what giant pandas really need[J]. Biology Letters, 7(3): 403-406.
[34] Zhao C, Hu J, Li YH, et al. 2014. Habitat use of Sichuan sika deer in forest, bush and meadows in the Tiebu Nature Reserve, Sichuan, China[J]. Pakistan Journal of Zoology, 46(4): 941-951.
[35] Zhao C, Yue BS, Ran JH, et al. 2017. Relationship between human disturbance and giant panda distribution in the Daxiangling Mountains[J]. Oryx, 51: 146-152.





