
�����Ǿƾ���֬����(non-alcoholicfattyliverdisease��NAFLD)�����˸������˵�һ���㷺�ļ����ס��ӵ�����֬�����Ե��Ǿƾ���֬���Ը���(non-alcoholicsteatohepatitis��NASH)�����ɽ�չΪ��Ӳ������ĩ�ڸβ�������NASH��ΪNAFLD�еĹؼ��Σ��о��䷢�����ƶ�NAFLD�����������൱��Ҫ�����á�
����
�����������о�֤ʵNAFLD�и�������������֬����Ĵ�лʧ�����ƣ������̴�����֢״̬�²���“�ڶ��δ��”���γ�NAFLD�Ļ����Բ��������ʵ���������Ȼ�̼(carbontetrachloride��CCl4)�����ζ���ʹС���ϸ������������Ӧ����ʹ�̴�����Ԫ����ϵ���(sterolregulatorybindingproteins��S��EBPs)�������ӣ����¸�ϸ����֬�ʼ��ۣ����ָ�ϸ����֬�����ԣ�ʵ��“��һ�δ��”�������ҵ����շ����Եͷ����֢�IJ�����̽����֢������Ϊһ��������Σ�������ڸ�ϸ��֬�����ԵĻ����Ϸ���“�ڶ��δ��”�γ�֬���εķ��ӻ��ơ�
����
����1 �����뷽��
����
����1��1 ʵ�鶯��
����
����6��8����C57BL/6J����С��(��������ҽ�ƴ�ѧʵ�鶯������)12ֻ��������20��26g������֤��:SCXK2012-0001������ҽ�ƴ�ѧʵ�鶯������SPF������������������ֱ�����С���Ϊ2�顣������(n=6):����Ƥ��ע����������40%��CCl4-ֲ������Һ(0��3mL/100g)2�ܺ��ٸ���Ƥ��ע��������ˮ0��5mL16��;��֢��(n=6):����Ƥ��ע����������40%��CCl4-ֲ������Һ(0��3mL/100g)2�ܺ��ٸ���Ƥ��ע��10%�ҵ���(casein)0��5mL16�ܡ�ʵ���������ȡС��Ѫ�������֯����80�汣�档
����
����1��2 Ѫ������֢���Ӳⶨ
����
��������������(serumamyloidA��SAA)��������������-α(tumornecrosisfactor-α��TNF-α)��Ũ��ʹ��ELISA�Լ��м��(�ֱ��������ң�D��˾������ʢ����Ƽ�����˾)��
����
����1��3 Ѫ��֬�ʺ����ⶨ
����
����С��Ѫ�����ܵ��̴�(totalcholesterol��TC)�����ܶ�֬�����̴�(low-densitylipoproteincholesterol��LDL-c)���ܶ�֬�����̴�(high-densitylipoproteincholesterol��HDL-c)Ũ�Ȳ���øż����ɫ�����(�Լ��й�������ٽ����\����˾)��
����
����1��4 ����HEȾɫ���ͺ�OȾɫ
����
����ȡ�������ʸ���֯��4%��ۼ�ȩ�̶����ֱ�����ʯ����Ƭ��������Ƭ��ʯ����Ƭ�ö��ױ��������ݶȾƾ���ˮ����ľ��Ⱦɫ8min������ˮ��ϴ����ȾҺ������Ⱦɫ2min��95%�ƾ���ˮ5min�����ױ���5min������������Ƭ��������Ƭ��10%������������Һ�̶�30min��1��2-����������2min���ͺ�O(Sigma��˾)Ⱦɫ10��15min����ľ����Ⱦ1min����ˮ��ϴ����10min�����ͷ�Ƭ���ٷֱ��������¹۲����֯֬������̶ȡ�
����
����1��5 ʵʱӫ�ⶨ��
����
����PC�Ҳ���Trizol����ȡ����֯�ܣ�NA����DEPCˮϡ�ͺ�ⶨŨ�ȼ�D(260)/D(280)ֵ(����ֵΪ1��8��2��0)���껯����ͬŨ�ȡ�������ת¼�Լ���(TaKa��a)����˵�����ܣ�NA��ת¼��cDNA������SYB��Greenӫ�ⶨ����T-PC�Ҽ�����֯�ĵ��ܶ�֬��������(low-densitylipoproteinreceptor��LDLr)���̴�����Ԫ����ϵ���-2(sterolregulatoryelementbindingprotein2��S��EBP-2)��S��EBP�ѽ⼤���(S��EBPcleavageactivatingprotein��SCAP)��m��NACtֵ���������м���1��ȡ2μL��ת¼�������ʵʱӫ�ⶨ��PC�ң���β-actin��Ϊ�ڲ��գ���Ӧ��ϵΪ20μL����������:95��30sԤ���ԣ�95��5s��58��30s��70��1min����40��ѭ����������Զ���ΔΔCt����2��С��������IJ��죬���㹫ʽ����:ʵ��������ڶ�����������ˮƽ�ı���=��exp(ΔΔCt)������ΔΔCt=ʵ����ΔCt��������ΔCt��ΔCt=Ŀ�Ļ���Ctֵ���ڲλ���Ctֵ��
����
����1��6 ������֯
����
������ѧȾɫ����֯�̶���ʯ��������Ƭ���ÿ�LDLrһ��(1��200����������ɭ)���ÿ�S��EBP-2һ��(1��300����������ɭ)���ÿ�SCAPһ��(1��250����������ɭ)4�������ҹ���ü����������黯�Լ���(������ɼ����)����Ⱦɫ�����岽�����˵���顣DABȾɫ���ݶȾƾ���ˮ�����ױ���������������Ƭ�������¹۲����࣬��ϸ�����ػ�ɫ�����ֲ�Ϊ���Ա��
����
����1��7 Westernblot���
����
�������ױ��ﰴ������ȡ�Լ���˵����ȡС�����֯�ܵ��ף���BCA�����е��������껯��ÿ����������Ϊ80��100μg��β-actin��Ϊ�ڲ��յ��ס���Ʒ�ӱ��Ի���Һ���10min��SDS-PAGE������Ӿ��Ȼ��ת��PVDFĤ��5%��֬�̷����·��2h���ֱ����һ��S��EBP-2(1��500����������ɭ)��һ��LDLr(1��200����������ɭ)��һ��SCAP(1��500����������ɭ)��һ��β-actin(1��5000��������ɼ����)��4���ҹ������TBSTƯϴ10min×3�Σ�����������������ø(H��P)��ǵĶ���(�������\������˾)��37�����2h��TBSTƯϴ10min×3�Σ�ECL�⻯ѧ��������ɫ��
����
����1��8 ͳ��ѧ����
����
��������SPSS17��0ͳ�����������н����x��±s��ʾ��2������ж�������t���顣
����
����2 ���
����
����2��1 ��֢���Ӽ����
����
������֢���SAA��(236��09±38��45)ng/mL�ݺ�TNF-α��(82.59±11��48)pg/mL�����Ը��ڶ�����۷ֱ�Ϊ(35��68±11��34)ng/mL��(17��91±2��77)pg/mL��P��0��05�ݡ�
����
����2��2 Ѫ��֬��ˮƽ�����
����
������֢���HDL-c��(0��91±0��10)mmol/L�ݡ�LDL-c��(2��65±0��44)mmol/L�ݡ�TC��(3��69±0��26)mmol/L�ݵ�Ѫ��Ũ������ڶ��������Խ��ͣ۷ֱ�Ϊ(1��80±0��12)��(5��76±0��33)��(4��16±0.22)mmol/L��P��0��05�ݡ�
����
����2��3 ����HEȾɫ���
����
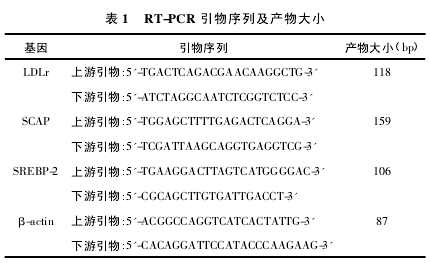
������ͼ1���Կ������������ϸ����ϸ������������״���У��ʷ���״�ֲ������뾲����Χ����֢������ṹ�ƻ�����ϸ���������ң��ɼ�����֬�ο��ݺ���֢ϸ������ÿ��ȡ3������֯��Ƭ��ÿ����Ƭȡ3��������Ұ������NAFLD���(NAFLDactivityscore��NAS)���֣����������֣�3�֣�Ϊ������ϸ����̬������֢��NAS���֣�4�֣������ΪNASH��
����
����2��4 С���ϸ���ͺ�OȾɫ���
����
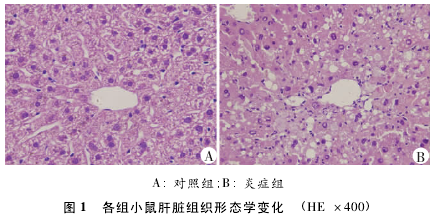
����ͼ2��ʾ������ɼ����Ƭ״��Ⱦ��������֢��ɼ�������Ⱦ�������۳�Ƭ����ʾ��֢�ɼ���CCl4������֬�����Ը�ϸ����֬�ʼ��ۡ�
����
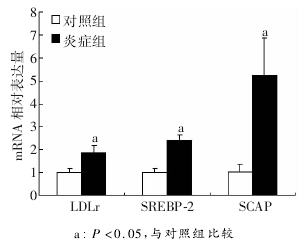
����2��5 С�����֯LDLr��S��EBP-2��SCAP��m��NA����ʵʱӫ�ⶨ��PC�Ҽ������ʾ����֢������ڶ������LDLr��S��EBP-2��SCAP����ı�����������(P��0��05��ͼ3)��

����
����2��6 ����֯�����黯Ⱦɫ���
����
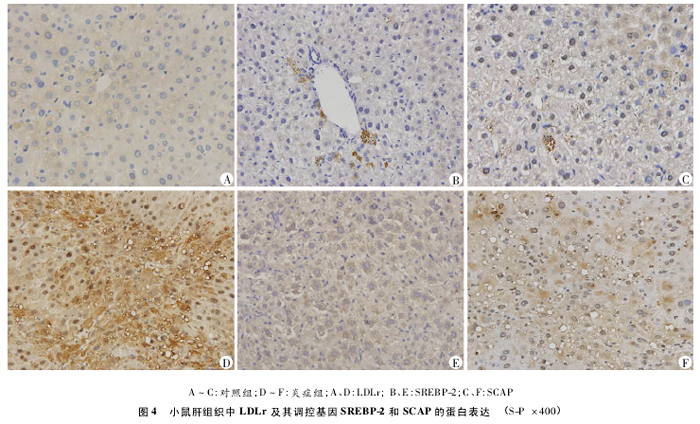
����S��EBP-2��SCAP���ױ����ڸ�ϸ���İ����У�LDLr���ױ����ڸ�ϸ���İ�Ĥ����ʡ���ԭ���Ա���Ϊ�ػ�ɫ��������ɫ����ͼ4�ɿ�������֢���LDLr��S��EBP-2��SCAP�ı���ȶ��������ࡣ
����
����2��7 Westernblot�����
����
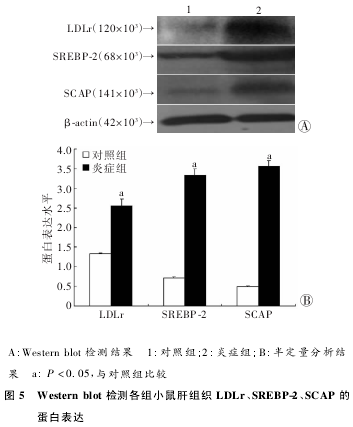
����Westernblot�������ʾ�����������ȣ���֢��LDLr��S��EBP-2��SCAP���ױ����������ߣ���m��NA��������(ͼ5)��
����
����3 ����
����
����NASH��ΪNAFLD��չ��һ���ؼ��㣬�䷢�����Ƹ��ӣ�Ŀǰ�����������ܵ���“���δ��ѧ˵”������������NAFLD��������������֬����Ĵ�лʧ���������д��������������ڵ��̴��Ĵ�лʧ�������о����١�
����
�������е��о����֣�����ά�ֵ��̴���̬ƽ��Ĵ�л����������3��������е���:ͨ��LDLr��ȡ�������̴�����ͨ��3-�ǻ�-3-���������øA��ԭø(3-Hydroxy-3-methylglutaryl-coenzymeAreductase��HMGC��)���ڵ��̴�����Դ�Ժϳɣ�������ĵ��̴���ͨ�����������ս�Ϻ�ת����A1��G1ת��ϸ��֮�⡣���л���ͨ��LDLr����Ѫ�����̴���ϸ���ڵ��̴�����Ҫ��Դ�����о�ͨ����������ʵ��֤����ϸ����“���δ��”�γ�����֬���εĵ��̴���лʧ�����ơ�����CCl4������ϸ��Ĥ֬�ʹ�������Ӧ������������Ӧ����������Ӧ��ʱ�����̴������ģ��Ӷ�����S��EBPs������SCAP����γɸ������������ת�Ƶ��߶����壬����������ø(S1P��S2P)�������£�S��EBPs���ѽ⣬�����ת¼���Ե�N-ĩ��Ƭ�ν���ϸ�����ڣ��ٽ�LDLr����������֬���ø�Ȱл�����յ�������Ӹ�ϸ�����̴��ļ��ۣ��ƻ�������LDLr-SCAP-S��EBP-2�������ڣ���“��һ�δ��”;�ٽ����ҵ���Ƥ��ע�䣬ʹ������֬�����Ե�С���ڳ����Եͷ����֢״̬������ϸ�������֬��������ʧ����ϸ��ͨ��LDLr�쳣��������ȡѪҺѭ���еĵ��̴������̴���ת�˼��٣����¸�ϸ���ڵ��̴��������ۣ�ʵ�ֶԸ�ϸ����“�ڶ��δ��”��ʹ��ϸ���ɵ�����֬������ת��Ϊ���ص�֬���Ρ�
����
�������о���C57BL/6JС��2��CCl4ע����پ�16�ܵ��ҵ���ע�䴦������֢���SAA��TNF-αŨ�ȱȶ�������������(P��0��05)��˵��С������ϵͳ����֢ģ�����ɹ���HEȾɫ���ͺ�OȾɫ�У���֢��ɼ�������֬�����ݼ���Ⱦ��������ʾ��֢�ɴٽ�С���ϸ���е�֬�ʼ��ۡ���֢��С��Ѫ���е�HDL-c��LDL-c��TC�����϶��������Խ���(P��0��05)��˵����֢�ɴٽ���ϸ��ͨ��LDLr�쳣��������ȡѪҺ�еĵ��̴������ٽ���ϸ���ڵ�֬�ʼ��ۡ���T-PC�Ҽ������ʾ��֢���LDLr��SCAP��S��EBP-2��m��NA����϶�����ֱ�������1��87��5��24��2��39�����������黯��Westernblot���ɿ�����֢����3�ֻ���ĵ��ױ���϶������������ߣ�������������һ�£����������֢���ӿ���ͨ���ٽ�LDLr-S��EBP-2-SCAP�ı����ǿ��ϸ�����̴����쳣���룬���¸�ϸ��֬�ʵ��쳣���ۡ�
����
�����������������о���CCl4���µĸ�ϸ��֬�����������֬�ʴ�л��S��EBPs��ϵ������ͨ������ʵ��֤ʵ����֢������CCl4���µĵ�����֬���������“�ڶ��δ��”������SCAP-S��EBP2-LDLr�������������ڣ���ʹ��ϸ���쳣��������ȡ���̴���������֢������֬�����γɹ����е���Ҫ���ã�Ϊ֬���ο������Ƶı�Ҫ���ṩ�������ݣ�ͬʱҲΪNAFLD���Ƶ���ҩ�����ṩ����;������NAFLD���̴����쳣���۵Ļ��Ƹ��ӣ����������лʧ�������ͨ·���д���һ���о���





